
12.8 The Coprinopsis fruit body: making stems
Fruit body stems are not as clearly divided into tissues as are caps but they do show a level of differentiation and a distinct morphogenesis. Corner (1932) introduced hyphal analysis as a procedure to describe tissues of polypore fruit bodies and this is used in routine classification (CLICK HERE to view our ‘Hyphal differentiation’ Resources Box we introduced in Chapter 9). Over the years a number of different types of hyphae and a range of tissue types have been described (Corner, 1966). Corner coined the terms monomitic, dimitic and trimitic to describe tissues consisting of one, two or three kinds of hyphae, and hyphae in these different categories have been referred to as generative (because they ultimately give rise to the basidia and directly or indirectly to all other structures), skeletal (with thick walls and narrow lumen, but lacking branching and septation) or binding hyphae (which have limited growth and irregular often repeated branching). Corner also introduced the terms sarcodimitic and sarcotrimitic to describe fruit bodies that have two or three types of hyphae (respectively) of which one is inflated and with thickened walls.
All these, remember, were first defined in polypore fruit bodies, but Redhead (1987) recognised such structures in a group of gilled fungi that includes the general Agaricus, Collybia, Marasmius and Flammulina, so cellular differentiation in fruit body tissues is widespread. Indeed Fayod (1889) had already described the presence of narrow hyphae amongst the more easily seen cells of the fruit body tissues he examined. These were called ‘fundamental hyphae’ but they have not been used in identification in the way that the mitic system has in polypores; today we might call them something like ‘embryonic stem cells’, being undifferentiated vegetative cells able to differentiate into a diverse range of specialised cell types. Although there are many reports of the occurrence of differentiated hyphal cells in various parts of fungal fruit bodies it is the relative sizes of differentiated hyphal populations, their distributions and the way the populations change during development that are crucial to understanding cell and tissue functions and there is only one clear account of a quantitative hyphal analysis, and this deals with the stem of Coprinopsis cinerea (Hammad, Watling & Moore, 1993a).
In this study 5 µm thick transverse sections were cut from fruit body stems of a range of developmental ages. Hyphal profiles were counted and cross-sectional areas measured using image analysis software. For each section of each piece of stem examined, the area of every cell within two randomly chosen radial transects was measured; transects were 12 µm wide. Individual cells were measured in strict order through the transect, starting from the exterior of the stem and ending at its centre, or at the lumen in older stems (Fig. 22). Two distinct populations of hyphae were identified and categorised as narrow hyphae, with cross-sectional area less than 20 µm2, and inflated hyphae, with cross-sectional area greater than 20 µm2. These populations of hyphae may be randomly distributed, evenly distributed or clumped together. In a random distribution the presence of one individual does not affect the probability of another occurring near by; in an even distribution that probability is lowered while in a clumped distribution it is raised. Statistical analysis of spatial distributions of the two populations of hyphae showed that inflated hyphae were very strongly evenly distributed, regardless of the age of the fruit body (from 27 mm tall to 70 mm tall) and regardless of position within the stem. The even (that is, non-random) distribution of inflated hyphae implies some form of organisational control. On the other hand, the spatial distribution of narrow hyphae was essentially random.
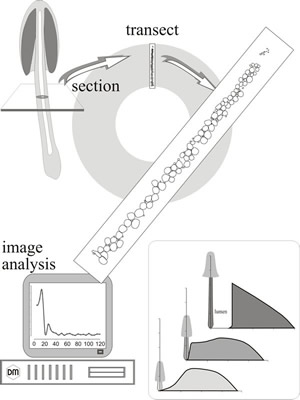
Both narrow and inflated hyphae can be seen in primordia only 3 mm tall. However, during stem elongation, the numerical proportion of narrow hyphae decreased implying that at least some (approx. 25%) of them are recruited to the inflated category. Low magnification images of transverse sections of any stem more than a few mm tall were dominated by highly inflated cells but narrow hyphae always constituted a significant numerical proportion (23% to 54%) of the cells in transverse sections, although, being 'narrow', they only contributed 1% to 4% of the overall cross-sectional area. Narrow hyphae stained densely with several conventional histological stains (especially Mayer’s haemalum, toluidine blue and aniline blue/safranin; all of which can stain nucleic acid/protein and protein/polysaccharide complexes) and revealed especially strong, particulate, staining with the periodic acid-Schiff reagent for polysaccharide.
However, not all narrow hyphal profiles in a transverse section and not all hyphal compartments belonging to any one narrow hypha in longitudinal sections stained equally. This differential staining might reflect differential function among the narrow hyphal population or, since narrow hyphae may be important in translocation of nutrients through the stem, it may simply reflect non-uniform vertical distribution of cytoplasmic materials caught in the process of translocation. The narrow hyphae seem to form networks independent of the inflated hyphae; they were seen to be branched and to be fused laterally with other narrow hyphae but inflated hyphae were neither branched nor associated in networks.
Generally, narrow hyphae were interspersed with inflated hyphae across the full radius of all stems irrespective of position along the length of the stem and irrespective of the developmental age. However, there was a progressive change in the distribution of inflated hyphae during maturation. In 6 mm and 27 mm tall fruit bodies the inflated hyphae increased in cross-sectional area from the exterior surface to about halfway across the radius but then size declined towards the lumen. In the 45 mm tall fruit bodies the cross-sectional area of inflated hyphae increased steadily from the exterior to the lumen and this pattern was even more pronounced in 70 mm tall fruit bodies. These measurements imply that expansion of the stem is mainly due to increase in cross-sectional area of inflated hyphae in an annular region just over halfway across the radius; I wonder what makes those specific stem hyphae expand. The geometrical consequences of cell inflation being emphasised in a distinct annulus within the stem tissue would be (Fig. 23):
- first, that the central core would be torn apart (it is not expanding, but the tissue exterior to it is expanding so the centre must tear apart), leaving its constituent cells as a remnant around the inner wall of the lumen so created;
- second, that the tissues in the zones outside the expanding annulus would be stretched and reorganised (they are not expanding, but the tissue interior to them is expanding, so outer tissue must be stretched).
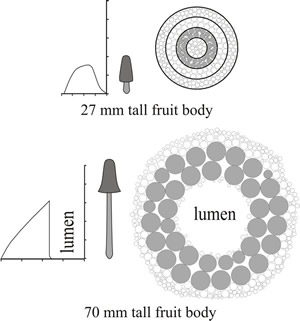 |
Fig. 23. Interpretation of the geometrical consequences of cell size changes during development of the stem of Coprinopsis cinerea. The two graphs show cell size distributions across the radius of the stem for 27 and 70 mm stems alongside scale drawings of the fruit bodies and diagrams of transverse sections of the stems, also drawn to scale. The diagrammatic transverse section of the stem of a 27 mm tall primordium (on left) is composed of solid tissue which is divided into four zones corresponding to the zones in the radius in the graph alongside. The central (zone 4) and outermost (zone 1) zones are comprised of rather smaller cells than the two cortical zones. During further growth the most dramatic cell inflation occurs in the cells of zone 3 which are here shown shaded. Growth from 27 mm to 70 mm in height is accompanied by a 3.6-fold increase in cell area in zone 3 but only 1.6-fold increase in zone 2. The cells in the other zones have to be rearranged to accommodate the inflation of zone 3 and a major consequence is that a lumen appears in the centre of the stem. Changes in cell profile area are shown to scale in these diagrams, though only a total of 327 cell profiles are illustrated. As this is only a tiny proportion of the cells involved in vivo, the diagrams inevitably distort the apparent relationship between cell sizes and stem size. Narrow hyphae have been ignored in these diagrams, though they are distributed randomly throughout the tissue and their conversion to inflated hyphae contributes to stem expansion. Adapted from Moore (1998a). |
What makes some hyphae become inflated while others remain morphologically similar to the vegetative mycelial hyphae is not known. However the organised pattern of inflation generates the final morphology of the mature stem. Cell inflation in a distinct annulus within the stem tissue turns a solid cylinder of hyphal tissue into a hollow tube, which itself is an engineering advantage as it produces a supporting element with greater stiffness per unit mass, and additionally produces another stretched-skin construction which gains even more structural strength from the combination of compression within the stem tissue and tension in its stretched outer tissues.
Updated July, 2019