Ericoid mycorrhizasThe order Ericales is a worldwide group of closely related families, perhaps best known in the UK by the Ericaceae family - heathland plants such as Calluna (heather), Vaccinium (bilberry) and Erica (heath). Heaths are wide open landscapes dominated by plants such as Heathers, Gorse and heathland grasses and punctuated by scattered trees such as Silver Birch (Figs 1 & 2). They are historic landscapes and are essentially a man-made habitat. Heathlands occur on barren infertile land. The soils are usually sandy (and therefore free-draining), acidic and very low in plant nutrients. Because the soils are free-draining, they do not hold water for long and heaths are therefore often subject to summer droughts. Under such conditions, fires are a constant hazard, particularly as much of the vegetation is very resinous and hence, inflammable. A unique association of plants and animals adapted to withstand such inhospitable conditions has evolved to form the distinctive heathland community.
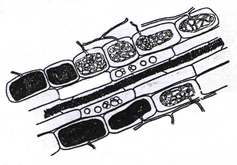
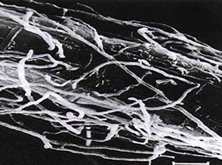
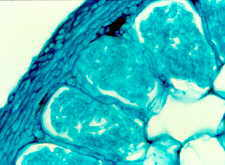
In the northern hemisphere, large areas of land can be dominated by the Ericaceae, whereas in the southern hemisphere, the Epacridaceae are most important in equivalent habitats. Heathland habitats are typically found at high altitudes and colder latitutudes, and have nutrient poor, acidic soils. It is likely that the success of the Ericales in this type of habitat is due to the endomycorrhizal associations they from; termed ericoid mycorrhizas. Some members of the order Ericales form arbutoid and monotropoid associations, which are dealt with in their own section. The fungi involved in ericoid associations are Ascomycota. The mycorrhizal roots formed by ericoid fungi show considerable uniformity through the Ericales. The primary roots of Calluna vulgaris, for example, grow vertically in the soil, giving rise to secondary roots, which terminate in very fine hair roots. It is the hair roots that are primarily colonised by ericoid fungi. The hair roots consist of an inner stele, surrounded by a cortex of layers; the outer exodermis and the inner endodermis. A single layer of epidermal cells forms the outer surface of the hair root (Figs 3 - 5).
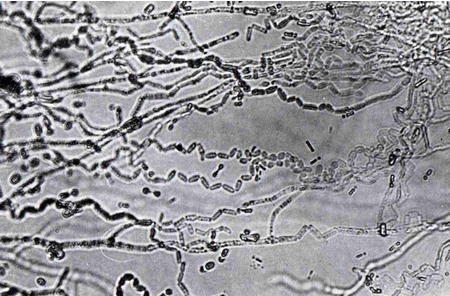
The ericoid fungal hyphae form a loose network over the hair root surface (Fig. 4); the hyphae can also penetrate the epidermal cells, often at several points in each cell (Fig. 3); and coiled hyphae fill the cell (Figs 3 & 5). Up to 80% of root volume can be fungal tissue and it is through these coils that nutrient exchange is thought to occur. Colonisation by ericoids is restricted to expanded epidermal cells (i.e. mature cells). Therefore the apical region of the hair root, behind the growing meristem, remains uncolonised until the cells differentiate and mature. The fungi involved in ericoid associations are facultative; that is, they can be free-living in soil, and can also be cultured in vitro. When grown on nutrient agar the fungi produce dark-coloured, slow growing, sterile mycelia. The absence of reproductive structures and spores has previously complicated the identification and classification of ericoid fungi. The segmentation of hyphae into zigzags (Fig. 6) by some species such as Hymenoscyphus ericae can help classify the fungi somewhat, but it is the increasing use of techniques examining DNA and RNA profiles that has advanced this area of research. It has been shown that there may be considerable genetic differences between isolates that are superficially similar in appearance.
Very little is known about the evolution of ericoid mycorrhizas. The association is geologically fairly recent, with fossil records of Ericaceae known from the early Tertiary (around 55 million years ago), and Epacridaceae from the Cretaceous. (To see a discussion of the evolution of mycorrhizas, click here). |


To skip to other mycorrhizal types, click below: |
|||
Close the window to return to your previous page |
|||