
6.2 Fundamentals of wall structure and function
Most cell walls are layered, and the innermost layer (that is, the layer immediately surrounding the plasma membrane) is a relatively conserved structural skeletal layer and the outer layers are more varied between species and are dynamically tailored to needs of the organism as it develops and matures and in response to interactions with the environment. Proteins rarely make up more than 20% of the wall material, and most are glycoproteins. Some proteins have a structural role but most contribute to the many other functions. Proteins at, or close to, the outer surface determine the surface properties of the wall; that is, whether it is hydrophobic or hydrophilic (= non-wettable or wettable), or adhesive, or antigenic. The low concentrations of lipids and waxes found in fungal walls usually serve to control water movement, especially to prevent desiccation.
Approximately 80% of the wall consists of polysaccharides. In hyphae a major component of the wall, and certainly the most important to structural integrity, is chitin, which is a β1,4-linked long polymer of N-acetylglucosamine (Fig. 1), the chemistry of which has been described in Chapter 5 (CLICK HERE to view the page). Chitin degradation, which is relevant to wall remodelling, is explored in Chapter 10 (CLICK HERE to view now).
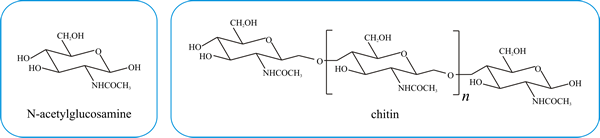 |
| Fig. 1. Covalent structures of N-acetylglucosamine and its linear homopolymer, chitin, which is synthesised by the enzyme chitin synthase. Natural sources of chitin have molecular masses of a few million; extraction processes fragment the polymer and the molecular mass of commercial preparations of chitin can vary between 350 000 and 650 000. |
Cell walls of filamentous fungi, such as Neurospora and Aspergillus, contain 10–20% chitin, but chitin accounts for only 1–2% of the yeast cell wall by dry weight (yeast chitin is mainly found in bud scars and only a very small fraction (around 2%) of the chitin content is found in the lateral walls). In both types of fungal cell, though, chitin molecules form into microfibrils, about 10 nm in diameter, by head-to-tail ‘crystallisation’ between the polymers. These crystalline polymers are held together by spontaneous hydrogen bonding and the microfibrils have enough tensile strength to provide the wall with its main structural integrity.
Chitin is a linear homopolymer chain but is frequently cross linked to other wall constituents, especially glucan (polymer of glucose) and mannan (polymer of mannose) polysaccharides. In almost all fungi, the main bulk component of the cell wall is a branched β1,3-glucan that is linked to chitin by a β1,4 linkage. Interchain β1,6 glucosidic linkages (the branch points in the branched glucan) account for 3% and 4% of the total glucan linkages, respectively, in Saccharomyces cerevisiae and Aspergillus fumigatus (Latgé, 2007). The β1,3 glucan is synthesised by a membrane-bound, GTP-stimulated, β1,3-glucan synthase. The wall may also contain some complex polysaccharides, like gluco-galacto-mannans.
The yeast cell wall is mainly composed of mannans, rather than glucans so we begin to see how the wall can contribute to taxonomy, because although there is a common structural theme, there are consistent differences in gross chemical composition of the wall that distinguish particular taxons. For example (see Gow et al. (2017) for references):
- the cell walls in the group traditionally known as zygomycetes contain chitin and chitosan (a polymer of glucosamine formed from chitin by removing the N-acetyl groups) amongst the more fibrous polymers, and polyglucoronic acid, and glucuronomannoproteins among the water-soluble gel-like polymers.
- Ascomycota and Basidiomycota both contain chitin together with the β1,3,
β1,6-linked glucans in fibrous form, but:
- Ascomycota feature a gel-like α1,3-glucan and galactomannoproteins. The yeasts Candida, Saccharomyces and the human pathogen Pneumocystis jiroveci have an outer cell wall containing highly mannosylated glycoproteins that covers the inner wall layers. In the human pathogen Histoplasma capsulatum, an outer wall layer of α1,3-glucan prevents the immune system recognising the invading cells.
- in Basidiomycota xylomannoproteins are found alongside the α1,3-glucan (Bowman & Free, 2006). The cell wall of the basidiomycetous yeast Cryptococcus is enveloped by a gelatinous capsule composed of glucuronoxylomannan (this makes up about 90% of the mass of the capsule) and galactoxylomannan that is anchored to the main wall via α1,3-glucan.
The taxonomic significance of the wall extends to the kingdom level. Not only are the walled cells of plants and fungi distinguished from one another by the use of cellulose as the main structural component in the former and chitin in the latter (compare Fig 1 above with Fig 1 in Chapter 10 (CLICK HERE to view the page), but the walls of hyphae of fungus-like members of kingdom Straminipila (Oomycota like Phytophthora and Pythium) contain cellulose microfibrils, which are about 12 nm in diameter, rather than chitin, alongside β1,3, β1,6-linked glucans. At the other end of the taxonomic spectrum, significant differences in the gross chemical composition of the walls of four commercial strains of the cultivated mushroom Agaricus bisporus have been detected (Calonje et al., 1995). Differences detected included overall composition of the wall and in the polysaccharide structures. In the face of such varietal differences, we have to be careful in assigning taxonomic significance to differences in wall structure.
Another feature that has been pointed out (Lesage & Bussey, 2006), is that the term ‘cell wall’ is applied to the extracellular matrices of fungi and plants, but an extracellular matrix is present in all three metazoan, fungal and plant kingdoms. Although the compositions of extracellular matrices do vary between taxonomic groups, all eukaryotic extracellular matrices result from the activity of a common cytoskeleton, which positions highly conserved secretory and construction machinery at, in, and through the plasma membrane.
Consequently, the evolutionary diversity that is observed seems to be only in the composition and structure of the extracellular matrices themselves, although this in itself must mean there will be some specialisation in the underlying cytoskeletal and secretory mechanisms involved in their construction (e.g. plant cells specialised to cellulose synthesis and assembly, fungal cells specialised to chitin synthesis and assembly, animal cells specialised to glycoprotein modification, etc.). On this view, then, cell walls are extracellular matrices specialised for physical containment that allow '...fungi and plants to build structures based on the use of cells as hydrostatic bricks...' (Lesage & Bussey, 2006).
The shape of the fungus is determined by its cell wall. The cell wall encloses the cell, providing osmotic and physical protection. The traditional view of the cell wall as a rigid structure, functioning primarily to withstand turgor pressure, has changed through time to a more dynamic view of the wall being a structure that is able to adapt to various conditions of growth, development and environmental stress, which, together with the plasma membrane and periplasmic space (the extracellular space between the plasmalemma and the wall), influences and regulates the flow of materials into and out of the cell. ‘Hydrostatic bricks’ might be an adequate description of plant cells, but fungal septa are incomplete (see the Cytokinesis and septation section of Chapter 5, CLICK HERE to view the page). Each fungal hypha is part of a closed hydraulic system which is under pressure. Osmotic influx of water occurs because of differences in water activity between the inside and the outside of the semipermeable plasma membrane.
The resultant tendency to increase cytoplasmic volume is counteracted by wall pressure due to the mechanical strength of the wall outside the plasma membrane. The difference between these two forces is the turgor pressure which is the resultant ‘inflation pressure’ keeping the hypha inflated. In a closed vessel the pressure is the same over the whole of the inside wall surface; this applies whatever the shape of the vessel. A fungal mycelium is a closed vessel divided up by septa which are perforated, and so, irrespective of its shape, the wall pressure is uniform throughout the mycelium.
The only way that pressure can be varied locally is to enclose part of the pre-existing mycelium by creating complete (= unperforated) septa or by plugging septal pores. This creates a second ‘pressure vessel’ (that might be a spore, haustorium, appressorium, or other specialised structure), which could then be provided with a different internal pressure to that of its originating mycelium. The pressurised mycelium or section of mycelium will remain intact for as long as the mechanical strength of its wall, everywhere over its entire surface, is sufficient to resist the internal pressure. The resulting turgor pressure has been estimated to be between 0.2 and 10 MPa (Money, 2004). ‘Typical’ atmospheric pressure at sea level is about 101 kilopascals (0.1 MPa) so this turgor pressure is equivalent to 2 to 20 times atmospheric pressure. Turgor pressure generates the force that enables hyphae to exert mechanical force on the substrates they are penetrating (Money, 2004; 2008) and these turgor pressures are sufficient to enable most vegetative hyphal tips to exert forces at least equivalent to atmospheric pressure at the micrometre scale of the hyphal apex. Of course, if the hyphal apex could be modified to a point, then even greater penetrating pressure could be exerted at the extreme tip. And this is exactly what happens in the appressoria (i.e. infection structures) of some plant pathogens such as Magnaporthe oryzae which have hyphal walls reinforced with melanin, making them the most robust of all walls found in nature and able to withstand an internal turgor pressure of up to 20 MPa and exerting forces many times atmospheric pressure at the nanometre scale of the appressorial apex.
In the context of the vegetative hypha, as long as the mechanical force that the wall can exert is equal to or greater than the force exerted by turgor, it will remain intact. However, if turgor exceeds the breaking strain of the wall at any point then the wall will rupture (possibly explosively locally), and the cell will die. This raises the problem we face in understanding apical hyphal growth; because the structure of the wall needs to be weakened to allow insertion of new wall material to permit the tip to elongate continuously and this must be achieved without exploding the tip (see our earlier discussion about growth at the hyphal apex (in the section entitled Cell biology of the hyphal apex in Section 5.16, CLICK HERE to view the page).
As long ago as 1976, Rosenberger compared the two-phase system of fungal walls (crystalline microfibrils embedded in an amorphous, gel-like matrix) with the two-phase systems of man-made building composites:
“Fungal walls contain a network of fibrils with the spaces in the net filled by matrix polymers and in this they resemble such man-made composites as glass-fibre reinforced plastic (GRP) and reinforced concrete." (Rosenberger, 1976).
This is a useful if not perfect analogy. Both systems are very strong for their weight, a property important for the ability of fungal walls to resist the turgor pressure of the cytoplasm. But we can best see how this analogy helps us understand the function of components of the fungal wall if we ask why civil engineers use this sort of building composite. In these building materials the matrix (the plastic in GRP or the cement plus aggregate in concrete) provides strength in compression and the reinforcing fibres (glass fibres in GRP or steel bars in concrete) improve strength in tension (stretching) and shear (force acting parallel to the surface) caused by bending. Applying this analogy to fungal wall shows that its ‘engineering’ produces a composite of highly cross linked glycan and polypeptides combined with chitin microfibrils (concentrated in wall layers nearest to the plasmalemma) that provides the strength overall to resist damage inflicted by compression, bending, and inflationary forces.
The civil engineering analogy can also be extended to tissues (see Chapter 12), but for the hyphal wall the shape and mechanical properties can presumably be altered, as they can in man-made composites, by changing fibre thickness, fibre length and fibre orientation. These changes bring about what we observe as hyphal wall rigidification and hyphal differentiation. We cannot leave this engineering analogy without bringing your attention to the growing interest attached to the use of the filamentous hyphae of mycelium cultivated in low-cost organic wastes to convert the wastes into useful and environmentally friendly materials. Although presently used primarily for a limited range of packaging and construction applications, a range of potential uses have been proposed for these mycelium composites, including acoustic dampers, super absorbents, paper, textiles, wound dressings and even electronic parts (Jones et al., 2017), as well as a range of building materials (Elsacker, 2021). .
In the true fungi, hyphal turgor is controlled to contribute to tip extension by driving the tip forward and shaping it by plastic deformation of newly-synthesised wall (Wessels, 1993; Bartnicki-Garcia et al., 2000). Chitin immediately provides the wall with most of its mechanical strength. If chitin synthesis is disrupted, or chitin is removed by experimental treatment with enzymes, the wall is sufficiently disrupted for the fungal cell to become osmotically unstable. We emphasise ‘in the true fungi’ here because members of the Oomycota like Achlya bisexualis and Saprolegnia ferax have little ability to regulate their turgor in response to external changes, probably because they grow only or predominantly in freshwater habitats, and they grow in the absence of turgor (Money & Harold, 1992, 1993; Lew et al., 2004); yet another difference between kingdoms Fungi and Straminipila, this one demonstrating that their superficially-similar filamentous growth forms are based on very different mechanisms.
The above is a highly generalised and abbreviated description of wall structure. Its purpose is to present a concept of the fungal wall at this point, rather than a specifically detailed constructional diagram. We will add detail to our description below, for the moment the concept is this:
- the main structural substance of the wall is provided by polysaccharides, mostly glucan, and in filamentous fungi the shape-determining component is chitin;
- the various polysaccharide components are linked together by hydrogen bonding and by covalent bonds;
- a variety of proteins and glycoproteins contribute to wall function, some of these are structural, some are enzymic, and some vary the biological and biophysical characteristics of the outer surface of the wall;
- proteins may be anchored in the plasma membrane, covalently bonded to wall polysaccharides or more loosely associated with the wall;
- the wall is a dynamic structure which is modified:
- as it matures, and/or
- as part of hyphal differentiation, and/or
- on a short-term basis to react to changes in physical and physiological conditions.
Finally, remember that, by definition, the wall is extracellular, it is a functionally-differentiated extracellular matrix and its entire structure lies outside the plasma membrane, so all additions to its structure must be externalised through the membrane before the wall can be restructured and the enzymic reactions that link wall components together are extracellular reactions carried out by secreted enzymes. In fact, fungal wall construction involves biosynthetic reactions that take place inside the cell in the endomembrane system, at the cell membrane by plasma membrane anchored enzymes, and in the cell wall itself. The major wall polysaccharides, chitin and glucan, are synthesised at the plasma membrane by transmembrane enzyme complexes that are delivered to the plasma membrane in an inactive form via secretory vesicles and then activated after insertion into the membrane (see below). In contrast, glycoconjugates, like the mannans, are synthesised intracellularly, in the endoplasmic reticulum and Golgi dictyosomes, where they may be conjugated to cell wall proteins, and then brought to the cell wall by secretory vesicles. Within the cell wall, polysaccharides and glycoconjugates can be crosslinked or branched by extracellular enzymes that reside within the cell wall.
Updated June, 2021