
6.7 Wall synthesis and remodelling
Newly synthesised chitin and glucan polysaccharides are either linear or shapeless molecules; they are cross-linked together and to other polymers to form the rigid three-dimensional network typical of the mature cell wall. The enzymes that accomplish this are called transglycosidases; these extracellular enzymes are either tethered to the plasma membrane with GPI anchors or located within the cell wall itself. The first cell wall-modifying transglycosidase to be recognised is a GPI-anchored enzyme encoded by the genes GEL and PHR in Aspergillus and Candida and GAS in Saccharomyces (Mouyna et al., 2013). This enzyme splits a β1,3-glucan molecule internally and transfers the newly generated reducing end to the nonreducing end of a different molecule; this elongates the glucan chain. Transglycosidases able to cross-link β1,6-glucan and chitin are also known (Arroyo et al., 2016); indeed, six families of conserved GPI proteins of this sort have so far been identified as being common across fungal species.
The transglycosidases are generally considered to be responsible for strengthening, or rigidifying, the fungal cell wall through the cross-links they generate. Cell wall hydrolases (like endo-β1,3-glucanase, and chitinase) and deacetylases, on the other hand, can plasticise the rigid cell wall. Chitinase is involved in bud separation in Saccharomyces, and endoglucanase is essential for cytokinesis in Schizosaccharomyces. In filamentous fungi, some models of apical wall synthesis suggest a delicate balance between hydrolysis of established polymers and synthesis of new wall material at the hyphal apex (see Section 5.16), although there is no clear evidence that cell wall hydrolases are required for hyphal tip extension. Deacetylases do appear to be widespread, and in zygomycetes, ascomycetes, and basidiomycetes substantial deacetylation of chitin (which produces chitosan) occurs. Chitosan is a more flexible molecule than chitin, and it is resistant to chitinases. Also, deacetylation of the chitin in the walls of plant pathogens can prevent the host plant’s receptors from recognising the pathogen and so delay the onset of plant defences.
During active hyphal growth wall construction activity is concentrated at the hyphal tip. Autoradiographic studies indicate that all synthesis of chitin and glucans takes place within 10 µm of the apex of the hypha of Neurospora crassa. The tip is highly plastic as the wall is laid down, but as walls mature they become more rigid. The rigidity is provided by the cross-linking of polymers to which reference is made above, thickening of fibrils and the deposition of materials in the interfibrillar matrix. The process is highly polarised, and reliant on maintenance of a positive turgor pressure within the cytoplasm (Bartnicki-Garcia et al., 2000). Wall rigidification in vegetative hyphae is a remarkably rapid process, taking only a couple of minutes. An estimate of the time elapsing between the deposition of plastic wall material at the hyphal tip and its subsequent rigidification at the base of the elongation zone (indicated by the point at which the hypha first attains a constant diameter) can be obtained by dividing the length of the tapered extension zone by the rate of hyphal elongation (Trinci, 1978).
Apical growth of the hypha requires long-distance transport between the subapical part and the apex of the tip cell. We have discussed some of the mechanisms that may be involved in Chapter 5. These mechanisms include cytoskeleton-based motors delivering vesicles containing enzymes and substrates over long distances to the hyphal tip (Bartnicki-Garcia, 2006; Riquelme et al., 2007; Steinberg, 2007, 2011). In addition, hyphal growth is accompanied by the secretion of exoenzymes that participate in both lysis and synthesis of fungal cell wall components which make the wall in apical regions flexible. Extension of the hypha can then occur as both turgor and cytoskeleton-based cytoplasmic expansion push the cytoplasm against the flexible apical wall. A summarised diagrammatic compilation of these events is shown in Fig. 4. This, together with the discussion in Section 5.16 should provide a sufficiently detailed view of fungal extension growth.
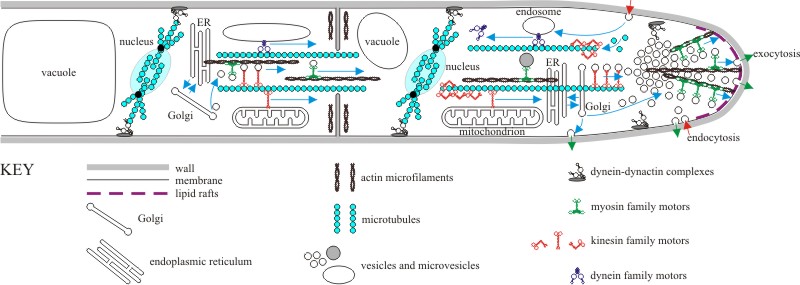 |
| Fig. 4. Cartoon representation of an overall molecular model of hyphal
growth. The key feature of hyphal apical growth is rapid movement towards
the apex of all the materials needed to create new wall, new membranes and
new cytoplasmic components. Most of these materials are exported in vesicles
by the endoplasmic reticulum (ER) and Golgi organelles, the vesicles being
delivered to the apical vesicle cluster (called the Spitzenkörper) along
microtubules powered by motor proteins of the kinesin and dynein families. The Spitzenkörper organises the final distribution of microvesicles along actin microfilaments to the plasma membrane at the extending tip. Vesicle fusion with the membrane is enabled by t-SNARE and v-SNARE proteins. Sterol-rich ‘lipid rafts’ at the hyphal tip could provide domains for apical proteins like signalling and binding complexes and might facilitate endocytosis. Endocytosis at the hyphal tip is dependent upon actin patches where myosin-1 polymerises actin into filaments that take the endocytotic vesicles away from the membrane. The extreme apex of hyphal tips undergoes extensive exocytosis, which is mainly devoted to synthesis of wall polymers outside the membrane and wall construction and maturation. Endocytosis features in the flanking regions of the hyphal tip, and this both recycles membrane components (originally delivered as exocytotic vesicles) and imports nutrients; both of which are transported to the endomembrane system for sorting and appropriate use. This figure also shows that (potentially many) subterminal hyphal cells contribute to the apical migration of resources; (streams of) vesicles, (trains of rapidly moving) vacuoles and mitochondria are all transported towards the apex and this transport extends through hyphal septa. Also note that the position of nuclear division spindles is probably specified by interaction between astral microtubules and membrane-bound dynein-dynactin complexes, and septal positioning is associated with rings of actin microfilaments. Remember: this IS a cartoon, no attempt is made to portray relative scale or relative timing (some structures, like division spindles) are more transient than others (like the Spitzenkörper). Also, everything happens, quickly; in the text we show that 38,000 vesicles have to fuse with the apical membrane each minute (that’s over 600 every second) to support extension of each hyphal tip of Neurospora crassa when it is growing at its maximum rate. See text of Chapters 5 and 6 for complete explanation, and refer to Steinberg (2007; 2011). |
We do not wish to over-emphasise apical wall growth here, because it is not the end of the story for the fungal wall. The picture that has emerged for the cell walls of yeasts and filamentous fungi alike is that the wall is an extremely dynamic construction with its components maintained in continual balance as wall enzymes repair and remodel the original wall. Cell wall damage in budding yeast, Saccharomyces cerevisiae, triggers a salvage mechanism called the cell-wall-integrity pathway consisting of at least 18 cell-wall-maintenance genes controlled by a single transcription factor and a specific signal transduction pathway. Sequences belonging to this integrity pathway are conserved in several yeasts and filamentous fungi (Bowman & Free, 2006; Klis et al., 2006; Gow et al., 2017).
We discuss the dynamic nature of the fungal cell wall elsewhere in this text in discussions of:
- hyphal and spore differentiation (CLICK HERE to view the page in Chapter 9),
- hyphal branching (CLICK HERE to view the page in Chapter 4),
- septation (CLICK HERE to view the page in Chapter 4; and CLICK HERE to view the page in Chapter 5),
- hyphal anastomosis (CLICK HERE to view the page in Chapter 5).
All these processes require that wall synthesis is restarted within a mature wall at a very closely-controlled place and at a specific time. Another relevant circumstance to be aware of is that two hyphal branches that must be joined together will synthesise a joint wall, and the resultant join can be stronger than the original hyphal walls (CLICK HERE to view the page in Chapter 12). All such remodelling depends on the coordinated activity of several glycoproteins already present within the wall structure. The ‘wall-associated enzymes’ involved include chitinases, glucanases and peptidases (Adams, 2004; Seidl, 2008; Gow et al., 2017); enzymes that hydrolyse and break down cell wall components, as well as glycosyltransferases which are involved in the synthesis and cross-linking of wall polymers. Only two chitinases of several detected in Coprinopsis cinerea, namely ChiE1 and ChiIII, hydrolyse insoluble crystalline chitin powder, while other C. cinerea chitinases do not, suggesting that crystalline chitin components of the cell wall (specifically in this study, in the mushroom stipe) are the targets for activity of ChiE1 and ChiIII (Zhou et al., 2019).
In addition to these instances of remodelling, there are many observations of secondary hyphal walls being synthesised as internal thickenings mostly made up of thick fibrils. These are probably glucans accumulated as intermediate to long-term nutritional reserves. The first detailed observations of this were made by Jos Wessels in the 1960s (reviewed by Bartnicki-Garcia, 1999) who showed that during the final stages of maturation of the Schizophyllum commune fruit body when its nutritional support is completely endogenous, requiring no external sources of nitrogen or carbon, an alkali-insoluble cell wall component, which was called R-glucan, was the main fraction of the wall to be broken down. R-glucan contained both β1,6 and β1,3 linkages, and was distinct from S-glucan which was alkali-soluble and constituted the bulk of the cell wall material left after mobilisation of the R-glucan.
Studies of the mobilisation process indicated that cell wall degradation correlated with cap development and was controlled by changes in the level of a specific R-glucanase enzyme. It seems that while glucose remains available in the medium carbohydrate is temporarily stored in the form of R-glucan in the walls of mycelial and fruit body hyphae. During this phase of net R-glucan synthesis the R-glucanase is repressed by glucose in the medium, but when this is exhausted the repression is lifted, R-glucanase is synthesised and by breaking down the R-glucan it provides substrate(s) specifically required for fruit body development.
The story is slightly different in Coprinopsis cinerea. In this organism glycogen seems to serve a similar sort of function to the R-glucan of Schizophyllum commune during fruit body development (glycogen is an α1,4 /α1,6 linked glucan (see Breakdown of polysaccharide: starch and glycogen section of Chapter 10, CLICK HERE to view the page); the reason may be that fruit bodies of C. cinerea develop much more rapidly than those of S. commune and the glycogen represents a much more efficient transient reserve that enables large quantities of sugar to be rapidly translocated through the fruit body with no disturbance to solute balance. Glycogen is involved in various aspects of vegetative morphogenesis in C. cinerea (Waters et al., 1975b; Jirjis & Moore, 1976), and the most recent transcriptome and sRNA sequencing have shown glycogen and trehalose to be the key energy sources during the outgrowth of basidiospore germ tubes in this organism (Lau et al., 2020), and clarified the environmental, physiological, and transcriptional aspects of carbon metabolism in all four developmental pathways of C. cinerea, namely vegetative mycelium, oidia, sclerotia and sporophores (Xie et al., 2020).
But wall glucans also contribute to vegetative morphogenesis in Coprinopsis cinerea. Mycelium of C. cinerea forms multicellular sclerotia, about 250 µm diameter, as resistant survival structures which pass through a period of dormancy before utilising their accumulated reserves to ‘germinate’ by producing a fresh mycelium. Glycogen is synthesised and accumulated in young sclerotia, but is not the long term storage product. For long term storage much of the carbohydrate is converted into a form of secondary wall material, probably glucan. Cells in the central bulk of the sclerotium may become extremely thick-walled, the primary walls being thickened on their inner surfaces by loosely-woven and very large fibrils, the development of which coincides with the gradual disappearance of glycogen from the cells (Waters et al., 1972, 1975a & b).
What we mean by ‘thick walled’ is illustrated in the electronmicrographs of Figs 5 to 9. Secondary wall is external to the plasma membrane but internal to primary wall. The illustrations in Figs 7 to 9 make it clear that the secondary walls can come to make up a very considerable proportion of the cell volume. This inevitably constricts the protoplasm to a correspondingly smaller central lumen, but the indications are that these cells remain alive; being effectively dormant until the sclerotium is presented with amenable growth conditions (Erental et al., 2008).
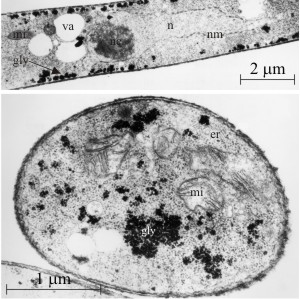 |
Figs 5 to 9. A selection of transmission electronmicrographs (TEMs) of submerged mycelium and sclerotia of Coprinopsis cinerea showing the two main types of thick walled hyphal cells; rind cells with dense, pigmented secondary walls of heavily-melanised glucan on outer and side walls which make up a plate-like layer of protective rind which resists environmental extremes, and the secondary walls of medullary cells which are uniformly thickened around the cell with large, branched fibres of glucan. This image, Fig. 5, shows a TEM of typical young vegetative hyphae from the submerged mycelium of Coprinopsis cinerea. Hyphae shown in longitudinal (top) and transverse sections. Note the thin (primary) hyphal walls that characterise this undifferentiated tissue. Key: n = nucleus, nc = nucleolus, nm = nuclear membrane, mi = mitochondrion, v = vacuole, er = endoplasmic reticulum, gly = glycogen granules. Electronmicrograph by Henry Waters. |
|---|
 |
 |
|---|---|
| Fig. 6. Secondarily-thickened walls of rind cells on the outside of sclerotia of C. cinerea. These secondary walls provide a protective layer and are heavily melanised. Electronmicrograph by Henry Waters. | Fig. 7. Secondarily-thickened walls of the central (medulla) region of a young sclerotium showing a cell with a secondarily-thickened wall in transverse section alongside many normal cells. This secondary wall is not melanised; it is constructed of thick fibres of glucan and is eventually recycled to provide carbohydrate resources when the sclerotium germinates. The granules (labelled gly) are accumulations of glycogen, which forms a short-term carbohydrate resource. Accumulation of glycogen occurs before formation of the glucan fibres and as the fibres are formed the glycogen content of the cytoplasm declines. Note the dolipore septum (ds) at top left. Electronmicrograph by Henry Waters. |
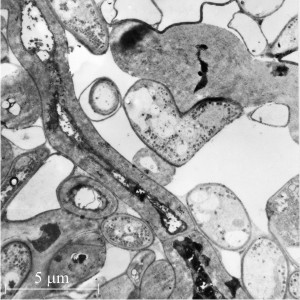 |
Fig. 8. Medullary cells with secondarily-thickened wall shown in longitudinal section, illustrating that the thickening (and consequent constriction of the cell lumen) is fairly uniform over the length of the hypha, and that the thickening can cross and involve the dolipore septa. Electronmicrograph by Henry Waters. |
|---|
 |
|---|
| Fig. 9. Magnified images of a longitudinal section of a secondarily-thickened wall showing its fibrillar structure. The region of secondarily-thickened (glucan) fibres highlighted in the left hand image is shown at extreme magnification on the right. Electronmicrographs by Henry Waters. |
Dormant sclerotia may survive for several years, being protected by a rind composed of tightly-packed hyphal tips which develop another form of secondary thickening which is a heavily-melanised thickened wall that forms an impervious surface layer (Fig. 6). Melanin is a dark coloured pigment (almost black at high concentration); it is a high molecular weight polymer of phenolic and/or indolic compounds. Phenols have hydroxyl group(s) bonded directly to aromatic hydrocarbons; indoles are heterocyclic organic compounds containing nitrogen, indole itself consists of a six-membered benzene ring fused to a five-membered nitrogen-containing pyrrole and has the formula C8H7N. These are negatively charged hydrophobic pigments that protect the hyphae and spores when they are cross linked into the cell wall structure. Normally, they are so completely interlinked, in fact, that it is possible in the laboratory to digest away all the other wall components to leave ‘melanin ghosts’ of the entire original cell wall (Dadachova et al., 2008). Natural melanins are biocompatible conductors. Nanocomposites using melanin nanoparticles extracted from squid inks have potential for use in bioelectronic devices such as biosensors and implantable devices (Eom et al. 2017). Fungal melanin ghosts, with their greater structural integrity, could have more interesting applications.
Melanin is extremely resistant to chemical and enzymic attack and contributes to virulence in many pathogenic fungi (for example Paracoccidioides brasiliensis, Sporothrix schenckii, Histoplasma capsulatum, Blastomyces dermatitidis, and Coccidioides posadasii) by reducing the susceptibility of melanised fungi to host defences and drugs (Taborda et al., 2008). Melanin increases cell wall rigidity, enabling hyphae of black fungi to penetrate host tissues and pigmented conidia or yeast cells to remain turgid when desiccated. Fungi with melanised walls are also resistant to electromagnetic and ionising radiations; the pigment seems to provide both physical shielding (from UV light) and quenching of cytotoxic free radicals (caused by ionising radiations). In fact, pigmented fungi may be resistant to the conditions of the surface of Mars, making them a contamination risk if inadvertently taken there by spacecraft missions. This survival potential in Mars-like conditions has been demonstrated in NASA balloon flights to the middle stratosphere (about 38 km altitude) of Earth, where radiation levels resemble values at the martian surface (Cortesão et al., 2021).
Although the detailed structure of melanin is unknown, two main types of melanin are found in the fungal cell wall, which are named after precursors in their biosynthetic pathways. DHN-melanin is synthesised from 1,8- dihydroxynaphthalene; and DOPA-melanin, which is synthesised from 3,4-dihydroxyphenylalanine (L-DOPA). In various fungi, there is evidence that melanin can be cross linked to mannans, mannosylated proteins, galactoxylomannan, chitin or chitosan (Gow et al., 2017).
Other cell wall pigments, the carotenoids (Chapter 10, CLICK HERE to view page), also protect against UV radiation. In general, mutants of the entomopathogenic fungus Metarhizium anisopliae with white conidia are more sensitive to UV radiation than mutants with purple conidia, which were more sensitive than mutants with yellow conidia, which in turn were more sensitive than the green wild strain (Braga et al., 2006). Wall pigments have a function; they’re not just pretty colours.
Updated May, 2021