
Evolution of mycorrhizas
Life first emerged on land during the pre-cambrian period (see Fig. 5 below for the geological timescale), when it was colonised by phototrophic micro-organisms, which were probably prokaryotic. Until fairly recently, it was believed that land plants became established during the late Silurian. However, recent evidence from several disciplines suggest that they may have emerged earlier, during the Ordovician period.
Vascular plants almost certainly arose from green algae, which become semi-aquatic, and then fully terrestrial during their evolution into the first land plants. When semi-aquatic algae did emerge and begin to invade the land, between 490 and 409 million years ago (Mya), they encountered a harsh environment. The soils contained no organic matter and therefore only mineral nutrients. This poor soil also meant that any nutrients or water obtained were quickly lost.
However, the barren land was also a place of opportunities, since there was very little competition, and carbon dioxide and light were readily available. To exploit this new environment, the earliest plants could either develop their own means of gaining nutrients and water or, as most did, they could alter their relationship with the aquatic fungi that were also invading the land. The earlist fungi were aquatic; chytrids with a small simple thallus (virtually a single cell), rhizoids to anchor the thallus to the substratum and extract nutrients, and flagellated motile spores.
Neither the alga nor the fungus was fully equipped to exploit the terrestrial environment. The alga lacked the ability to extract essential nutrients from the soil (what there was of it), whilst the fungi lacked the ability to manufacture carbohydrates. Both had problems dealing with desiccation.
Instead of the fungi becoming parasitic on the rhizoids of the evolving plants (roots or root hairs had not yet evolved), the two organisms formed a mutualistic symbiosis that allowed them both to exploit the terrestrial environment.
The algal/plant component became the morphologically dominant partner, as it was better equipped to survive terrestrially due to autotrophy, and hence the plant component became the rapidly evolving partner. The partnership, (perhaps representing the earliest lichen) was capable of more efficient uptake and assimilation of nutrients, and this could also include nutrients derived by the fungal partner's ability to digest and recycle the two-billion year accumulation of dead bacterial remains washed up on the shores of the shallow seas and lakes. Thus, the partnership would have had a selective advantage over non-symbiotic plant forms, not only for survival, but also for dynamic evolution, since the extra energy generated using extra nutrients would allow for more differentiation and development of complex tissues.
The fungal thalli involved in these early associations almost certainly gave rise to the aseptate hyphae (which we now classify among the Zygomycota), although whether this was before or after the emergence of land plants is unknown. However, as most microbes today thrive best in biofilm communities it is a fair assumption that this was also true of the most ancient microbial communities. Life in a biofilm might have been adequate for a semi-aquatic/semi-terrestrial partnership between unicellular fungi and unicellular algae. But the filamentous mode of growth is an excellent way of escaping the biofilm to explore dry land. Fossil and DNA evidence suggests that zygomycete aseptate hyphae originated during the Cambrian, and resemble those fungi that form arbuscular mycorrhizal associations today (Figs 1-3). The septate fungi (Ascomycotina and Basidiomycotina) do not emerge into the fossil record until the Devonian period.
 |
 |
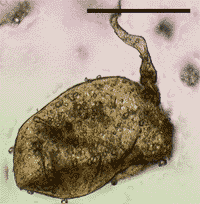
| Fig. 1. Light micrograph of a fossil fungal spore from the Ordovician period, 460 Mya. Reprinted with permission from Redecker, D., Kodner, R. & Graham, L.E. (2000). Glomalean fungi from the Ordovician. Science, 289: 1920-1921. © 2000 American Association for the Advancement of Science with permission. | Fig. 2. Modern-looking 'vesicles' in the 'roots' of a Devonian plant fossil. From Kendrick (1999) The Fifth Kingdom. © Mycologue Publications with permission. |
The AM fungi are one of the few plant-fungus relationships that have a fossil record (Simon et al, 1993) and it is generally accepted that early vascular plants were associated with AM-like fungi, and that their origin and ability to colonise land was highly dependant upon the association (Lewis, 1987; Selosse & LeTacon, 1998; Allen, 1991).
Ribosomal DNA sequencing by Simon et al (1993) place the origin of AM-like fungi between 462 and 363 Mya, within the Ordovician, Silurian and Devonian periods. These dates would easily place them at the time of land plant emergence.
Fossil mycorrhizas were first discovered by Weiss (1904) in lower carboniferous strata. The earliest and best examples of endomycorrhizas are from the Rhynie Chert fossils, from the Devonian period, discovered by Kidstone & Lang (1921). These show fungal structures resembling vesicles and spores, from the fungus Palaeomyces, associated with the rhizoids of plants such as Rhynia and Asteroxylon.
Recent reappraisal of the Rhynie Chert plants suggest those primitive plants may have been associated with fungi very similar to modern AM fungi by the early Devonian period (410 to 360 Mya) (Pirozynski & Dalpé, 1989).
The most important recent discoveries of mycorrhizal fossils are the arbuscles found by Stubblefield et al (1987) in Triassic strata in Antartica. These fossils show arbuscles organised like those of modern day AM fungi, indicating that the structural characteristics of the AM were well developed by the Triassic, even if the functional properties may not have been.
 |
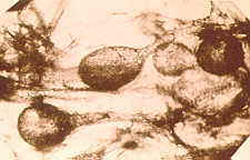
| Fig. 3. Fossil arbuscular mycorrhizas in Aglaophyton from the early Devonian. From Taylor et al., (1995) Fossil arbuscular mycorrhizae from the Early Devonian. Mycologia, 87: 560-573, with permission. |
The AM association is now widespread and found in most of the extant plant families. Its worldwide dominance is such that the association is now regarded as ancestral in plants (see Selosse & LeTacon, 1998).
Other mycorrhizal forms evolved from the AM-like at varying rates. The evolution of ectomycorrhizas (ECM) is a relatively recent event geologically, with a fossil record dating back to the second half of the Mesozoic. This is consistent with the emergence of its associated plant taxa. Pinaceae may have originated by the late Triassic, and certainly by the Jurassic, with Pinus in existence by the start of the Cretaceous (Fig. 5). However, the main differentiation of the family occurred from the mid-Cretaceous onwards, when Betulaceae, Fagaceae and Salicaceae originated (Malloch, D.W., Pirozynski, K.A. & Raven, P.H. (1980) Ecological and evolutionary significance of mycorrhizal symbioses in vascular plants (A Review). Proceedings of the National Academy of Sciences USA, 77: 2113-2118). The fungi associated with ECM, septate Ascomycotina and Basidiomycotina were certainly present during the Cretaceous (130 Mya) and ascomycotina possibly earlier.
Orchid and ericoid mycorrhizas most likely arose early in the evolution of their host taxon, but are more difficult to date. The ericoid association is geologically fairly recent, with fossil records of the Ericaceae dating back only to the early Tertiary.
 |
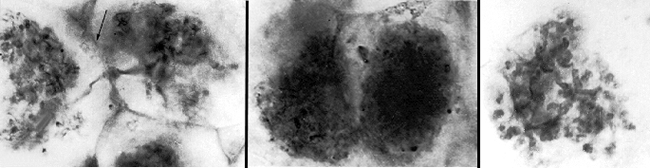
Fig. 4. Possible orchid fossil (Protorchis monorchis) from the Eocene epoch of the Tertiary period, 54 Mya. From Arditti (1977). Orchid Biology: Reviews and Perspectives I. Cornell University Press, London. © Arditti with permission. |
The Orchidaceae probably arose before the ericoid, during the Eocene, a division of the Tertiary period beginning 54 Mya. Several orchids or proto-orchids have been identified from this period, such as the fossil above, although whether these are true orchids is still debated. Fossils evidence is scarce though, since orchids do not fossilise well (Fig. 4).
 |
|---|
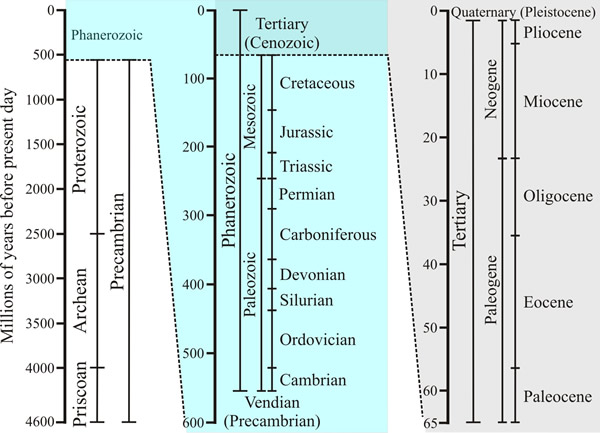
| Fig. 5. The geological timescale. The figure shows how the entire span of time during which the Earth has existed is classified into eras and epochs. Time axes on the central and right-hand panels are magnified views of the upper section of the previous panel. The last 65 million years is called the Cenozoic era, and is divided into a number of epochs, which began with the Paleocene approximately 65 million years ago, and runs to the present day. The Paleocene, which lasted from 65 to 54 million years ago, is followed by the Eocene (54-34 million years ago), the Oligocene (34-24 million years ago), the Miocene (24-5 million years ago), the Pliocene (5-1.8 million years ago) and the Pleistocene or Ice Ages (1.8 million years ago to 10,000 years ago). The period since the last retreat of the glaciers and present glacial warming is called the Holocene, or Recent (10,000 years ago to present day). Source: http://www.geo.ucalgary.ca/~macrae/timescale/timescale.html. Figure and caption from Moore et al., 2011 (URL). |
Further information about mycorrhizas and the evolution of fungi and the ecosystems to which they contribute can be found in the new textbook 21st Century Guidebook to Fungi by David Moore, Geoffrey D. Robson & Anthony P.J. Trinci. Published 2011 by Cambridge University Press: ISBN: 9780521186957. URL: ttp://www.cambridge.org/gb/knowledge/isbn/item6026594/?site_locale=en_GB. View Amazon page.
Updated December 15, 2016