解讀真菌的形態發育
趙紹惠 和 大衛摩亞
Department of Biology, The Chinese University of Hong Kong, Shatin, N. T., Hong Kong SAR, China
中國香港新界沙田 香港中文大學生物系
&
School of Biological Sciences, University of Manchester, Manchester M13 9PT, U. K.
英國曼徹斯特市 曼徹斯特大學生物科學學院
第三課:真菌之生物化學
真菌有獨特的營養吸收形式,首步是 : 分泌將環境中的聚營養物降解,釋放簡單的糖、氨基酸、炭酸、 嘌呤、嘧啶等的可吸收形式,明顯地,真菌在這方面有助物質的循環 (cycling) 和礦化 (mineralization)。
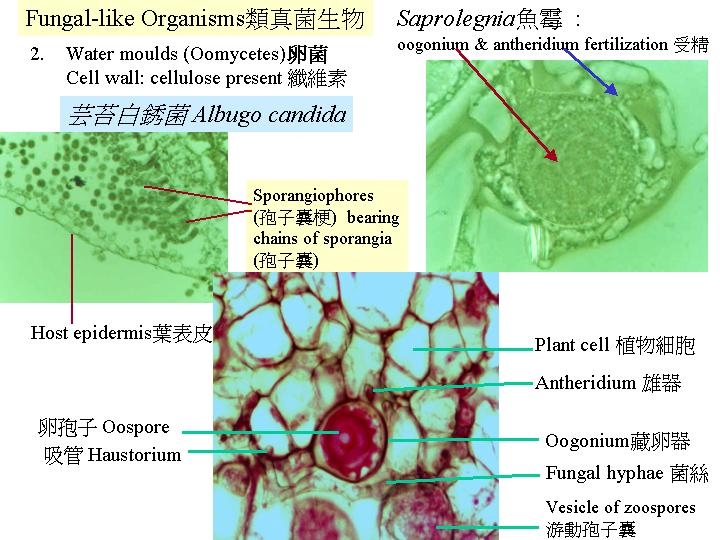
真菌有三類營養模式:最多的是腐生營養模式 (saprotroph) ; 真菌消化已死的有機生物 (Coprinus on horse
dung) 。腐食/腐屍性的真菌 (necrotroph) 會先將寄主組織殺死,然後吸收營養。相反,活食的真菌 (biotroph) 就是以活生物為食或寄生,而後者對寄主只引起局部的破壞,真菌菌絲長進寄主的內組織進行分解、吸收營養 (Albugo.jpg)。這類活食的真菌通常有專一的寄主 , 但腐生和腐食性真菌卻利甪不同的聚合體營養 (土壤、枯枝落葉層、木頭、和草食動物的糞便等),因此,首二類真菌擁有廣闊的棲息地 。
底物降解
植物生物量是由木質素 (lignin)、多糖、纖維素、半纖維素和膠質組成,其成分因應細胞和年齡而改變。還有這些成分不是單獨的而是組合成纖維木質素或木質蛋白等複合物。纖維素是地球上最豐富的有機化合物 ,佔有機炭物的50%或以上;大約每年有1,011 噸的合成,它是一種多糖。多糖是單糖透過糖鍵聚合而成。不同的糖數目和種類,再加上不同的鍵連接可產生各式樣的多糖。
類真菌的水霉的細胞壁也有纖維素,但纖維素是植物細胞壁的主要成分。纖維素是一沒分枝、單由葡萄糖以 beta-1→ 4連合而成的聚合體,分子量可由 50,000 到接近百萬不等。 纖維素的崩解化學上是直接的, 但因其複雜的物理形態,實際的崩解過程仍未完全了解。 纖維素在溫和酸水解下,釋放可溶性糖,和餘下有 100 - 300 葡萄糖長度的低聚糖殘留物,前者因可被崩解而被稱謂無定形纖維素 (amorphous cellulose),後者對酸水解有抗性的便是:結晶纖維素 (crystalline cellulose)。
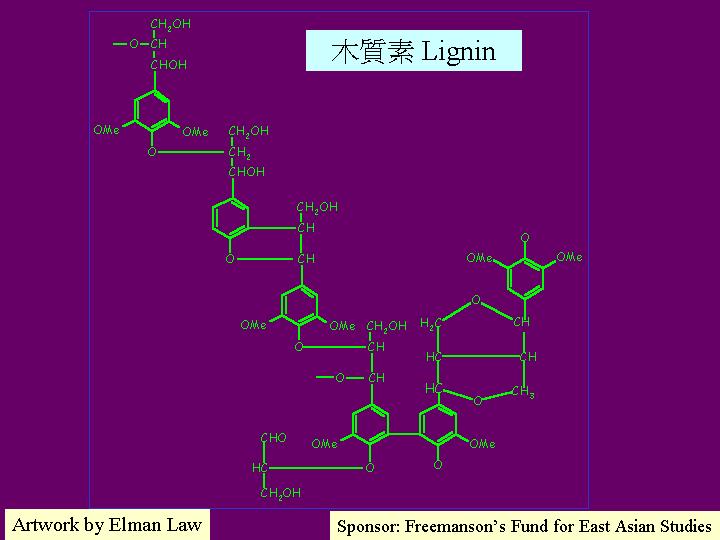
木質素(lignin) 是一高分子量、不溶性的聚合體,其構造複雜和多樣化,但大致上是由許多O-甲基化的苯衍生物組成的,尤以:松柏基醇 (coniferyl alcohol)、 芥基醇 (sinapyl alcohol) 和香豆基醇 (p-coumaryl alcohol) 為主,其相對比例在被子植物和裸子植物之間是不一致的。木質素本身極難生物降解,固能保護其他植物聚合體。少數微生物擁有降解木質素的能力,其中包括一些擔子真菌、一些子囊菌和一些細菌。眾多研究均認為木質素的降解不是作用,因為其複雜和多樣的結構與的專一性不附;木質素的降解是靠活性氧 (氧自由基)引致的,而過程就像燃燒作用,一觸發就擴散開去 (Reid, 1995);降解過程第一步驟是由一電子的氧化作用而產生不穩定的中間物,再引致廣泛而連串的氧化反應 (Kirk et al., 1990;
Hatakka, 1994; Cullen & Kersten, 1996)。不同真菌的木質素降解系統可有異同處 (diverse
enzymes)。
黃孢亮毛伏革菌 (Phanerochaete
chrysosporium) 能夠完全將木質素礦物化成 CO2 和水。它生產木質素降解所需的體外系統,包括:蟲漆 (laccase)、木質素過氧物 (lignin peroxidases, ligninase)
和錳過氧物
(manganese peroxidases, MnP)。黃孢亮毛伏革菌所生產的十五種木質素過氧物是一類糖化的含鐵蛋白 (heme protein) ,估計活性氧是結合在木質素過氧物的活性部位的,其分子量介乎 38,000 至 43,000之間,不同菌株和一菌株因應環境需求可生產不同的同功 (isozymes)。而黃孢亮毛伏革菌在停止生長後(即表現次生新陳代謝) 或氮飢餓誘導下,才生產木質素降解。不是每一真菌在低濃度氮下,才產生高木質素降解 (Chiu et al., 1998)。
木質素過氧物能氧化還電勢 (redox potential) 超1.4 V的二甲氧基的芳族化合物 (dimethoxylated aromatics) ,產生自由基作降解底物的機制,中間的氧化還原媒體 (redox active mediators) ,包括:真菌代謝物:藜蘆基醇(VA, veratryl alcohol)。
木質素過氧物利用過氧化氫的單電子氧化作用,產生芳基陽離子自由基 (aryl cation radicals) ,去切斷C-C 和 C-O 鏈,故表現底物非專一的特性,不像作用。既然外加的過氧化氫 (catalase) 破壞過氧化氫成氧和水後,木質素的降解就受到完全抑制;因此,木質素降解作用牽涉過氧化氫 (H2O2) 是得以肯定的。
真菌合成藜蘆基醇 (VA, veratryl alcohol) 、甲基酯 (methyl ester) 和苯甲醚是由靠氯甲 (chloromethane) 的甲基化系統 (chloromethane
(CH3Cl)-dependent methylation system) 負責的 (Harper et al., 1990) 。相比下,酚類底物 (phenolic substrate) 的甲基化作用由S-腺甲硫氨酸甲基化系統(S-adenosyl methionine (SAM) –
dependent methylation system) (即 3-O- and 4-O- 轉甲基(methyltransferase)) 負責的 (Jeffers et al., 1997)
。
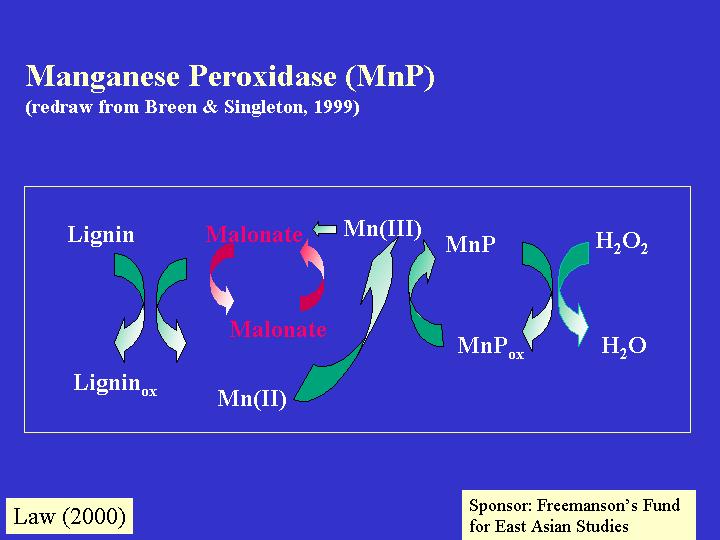
黃孢亮毛伏革菌的錳過氧化物則是第二類的含鐵蛋白 (Elisashvili, 1993;
Reddy & Dsouza, 1994; Broda et al., 1996; Cullen & Kersten,
1996)。MnP氧化 Mn2+至 Mn3+,在適合的真菌分泌的金屬螯合劑 (chelators) 穩定下,Mn3+就攻擊酚類底物和用脂肪過氧化作用 (lipid peroxidation) 去轉化非酚類底物 (nonphenolic structures) 。有些真菌就製造「LiP - MnP」雜種的過氧化物 (peroxidase) (Caramele et
al., 1999;Palma et al., 2000) ,如:側耳屬 (Pleurotus ergynii) 和煙管菌 (Bjerkandera) ,它們的氧化Mn2+ -過氧化對典型LiP 底物 (如:藜蘆基醇 (veratryl alcohol)) 有作用,擁有雙甲氧化 (dimethoxygenase) 和Mn2+ -無關連 (Mn2+
-independent) 的活性 (Böckle et al., 1999; Ruiz-Duenas
et al., 1999) 。
現在已知 Mn3+ 、藜蘆基醇 (veratryl
(3,4-dimethoxybenzyl) alcohol) 和活氧自由基 (activated oxygen species;e.g. HO., O2.-)
能氧化木質素單位。
H2O2可循岐化作用 (dismutation) 產生 (2 O2.- + 2H+ → H2O2
+ O2) 。
而Mn2+的氧化就伴隨Mn3+ 的生產 (O2.- + Mn2+ + 2H+
→ H2O2 + Mn3+)。
當然,HO. 的生產可循鐵催化的反應(iron-catalyzed Haber-Weiss
reaction) 而來 (O2.- + H2O2 → HO.
+ HO- + O2 ) 。
白色腐爛真菌的降解木質素系統中,氯化對甲氧基苯甲醇 (Chlorinated anisyl alcohols)
負起一重要的代謝任務;成為體外芳基醇 (aryl alcohol oxidases) 的可循環底物,因此,整個系統有H2O2的生產、還原 (quinones) 和含苯氧基的自由基 (phenoxy radicals) ,但防止木質素直接降解物的再聚合(repolymerization) (de Jong
et al., 1994) 。
生產H2O2的氧化包括:芳基醇 (AAO, aryl alcohol oxidase) 、乙二醛氧化 (glyoxal oxidase) 、葡萄糖氧化 (glucose oxidase) 和呲喃糖氧化 (六環糖氧化,pyranose oxidase) 等,都是降解木質素的關鍵。而H2O2在降解木質素時就負責接收木質素過氧化物釋放的電子 (electron acceptor) ,或作為反應物 (reactant) 去生產帶氧自由基,降解植物細胞壁。
利用免疫學的標簽法,再以掃描電子鏡和營光顯微鏡觀察,白色腐爛真菌(如:Pleurotus eryngii) 的芳基醇 (aryl-alcohol oxidase) 就被固定(immobilized) 在真菌分泌到體外的木質素-多糖複合物 (lignin-polysaccharide
complex,即菌絲鞘(hyphal
sheath)) 上 (Barrasa et al., 1998) 。因此,木質素的最初降解需要菌絲與底物的接觸。但是,Mn2+ 是微小的,可以擴散到底物中去,解除了直接接觸的限制,對底物作完全的降解。
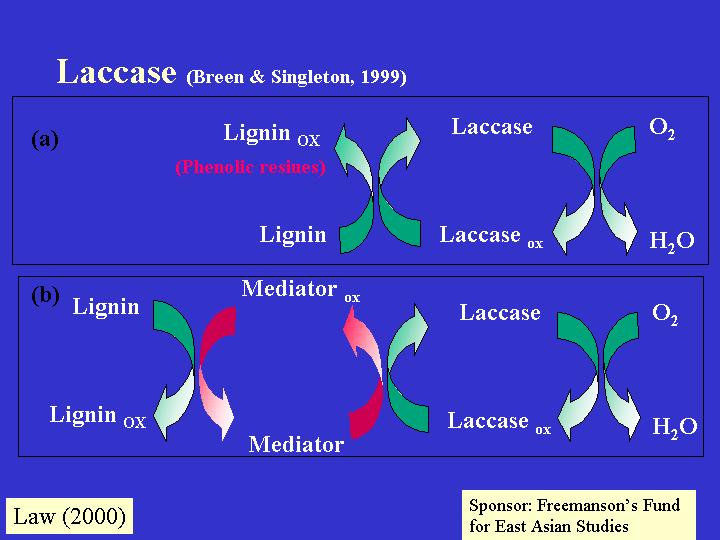
很少真菌製造木質素過氧化物,而是製造體外蟲漆 (laccase; EC 1.10.3.2) ,如:褐色腐爛真菌 (brown rot fungi)。蟲漆是五水的銅氧化,以氧為電子接受器去消除酚氫氧基團 (phenolic hydroxyl groups) 產生的氫自由基(hydrogen radicals) 。自由基製造後會進行重整,引發烷基-芳基的解離 (alkyl-aryl cleavage) 、基醇的氧化,和支鏈與芳環的解離 (cleavage of side chains and
aromatic rings) 。當適合的氧化還原中間媒體存在時,蟲漆更會氧化非酚底物。所以,褐色腐爛真菌也可製造蟲漆,它是含銅的加氧,可氧化 o- 和 p-酚 (phenols),並且負責木質素降解物的新陳代謝。雙孢磨菇 (Agaricus bisporus) 在菌絲生長期間,堆肥底物中的木質素被大量降解,也正是蟲漆的高峰期;蟲漆達蛋白質總含量的2% (Wood, 1980a)。錳過氧化物的活動與蟲漆的有相互關係 (Bonnen et al., 1994)。但當雙孢磨菇形成子實體時,蟲漆迅速地失活和被水解 (Wood, 1980b)。這些都反映真菌在進入生長發育過程時,不斷改變從環境所要求、提取營養。
白色腐爛真菌如黃孢亮毛伏革菌 (Phanerochaete chrysosporium) 和半知菌木霉 (Trichoderma reesei)
的纖維素由許多的水解所組成:內切葡聚糖、外切葡聚糖和纖維二糖 ( beta-糖) 協同增效和組成一複雜的體外纖維素顆粒 (Lemaire, 1996)。其中, 內切葡聚糖隨意攻擊纖維素, 產生葡萄糖、纖維二糖和一些纖維三糖。外切葡聚糖從纖維素分子的非還原末端開始攻擊,除去葡萄糖單位,或兼有纖維二糖水解的活性,從纖維素分子的非還原末端釋放纖維二糖。纖維素的結構包括:除接觸反應區外,還有許多觸媒作用、多種結合和底物附著的區域。這些區域將固定在底物上,集中水解的位置和方便複體增效,使分泌的個體/細胞容易吸收纖維素降解物:葡萄糖 (Beguin & Aubert, 1994; Radford
et al., 1996)。
白色腐爛真菌 (white rot fungus) 黃孢亮毛伏革菌分解纖維素時,會生產兩種纖維二糖氧化還原:纖維二糖: p-苯昆氧化還原 (CBQ;cellobiose: quinone oxidoreductase) 和纖維二糖氧化 (CBO; cellobiose oxidase)。 纖維二糖氧化 (CBO) 將纖維二糖氧化為 δ-內酯,同樣,纖維二糖:p-苯昆氧化還原 (CBQ) 也可產生δ -內酯,而 δ -內酯會最後被轉化成葡萄糖和葡萄糖酸。因此,這些纖維二糖氧化還原可能是管理纖維二糖和葡萄糖水平以免抑制葡聚糖內的活性。此外,纖維二糖氧化 (CBO) 還原Fe(III) 和連同過氧化氫 (H2O2) 作用下,產生氫氧基 (hydroxyl radicals) ,這些自由基就用來降解木質素。因此,推斷CBO可能連接纖維素和木質素的降解 (Ander, 1994)。不過, Eriksson et al. (1993)
有證據顯示:CBO/CBQ與木質素過氧化物 (lignin peroxidases) 和蟲漆 (laccases) 若同時存在,會減慢木質素降解物的氧化速率。
褐色腐爛真菌 (brown rot fungi) 則使用不同的首步驟去分解纖維素;縱然纖維素是被木質素聚合體所保護,褐色腐爛真菌分泌的過氧化氫和木頭裡的亞鐵離子 (Fe(II)) 去氧化、斷裂糖複合物,因而開放底物讓更多的水解作用。有提議:覆蓋在真菌菌絲表面的草酸鹽 (oxalate)會將木頭上鐵離子 (Fe(III)) 還原成Fe(II), 驅使纖維素的氧化崩解。白色腐爛真菌雖然生產木質素降解用的 H2O2,它們並不生產草酸鹽,因此,纖維素不被氧化分解。
另外,在有限葡萄糖和純氧大氣 (氧壓力,oxidant stress)下,白色腐爛真菌會製造木質素降解如:LiP,正因為 LiP消耗氧催化體外H2O2去成為水和自由基,後者更降解植物的木質素,暴露更多碳源。
澱粉是植物的主要儲備多糖,它是葡萄糖以 alpha- 1→ 4 糖鍵聚合而成。 直鏈澱粉是長的沒有分枝鏈的,然而澱粉糊精 (澱粉有75 - 85% 是以此形式存在) 則在 alpha –1 → 6糖鍵處有分枝。澱粉降解包括: alpha- 澱粉, 它們是內切, 只針對alpha- 1 → 4 糖鍵而不是alpha –1 → 6糖鍵;beta- 澱粉則是外切, 將直鏈上的隔跳1,4鍵切斷直至到1,6鍵的分枝點, 產雙糖麥芽糖 (maltose);氨基糖 (或 葡萄糖澱粉)對1,4鍵和1,6鍵都起作用,它像是真菌獨有的;脫支將1,6鍵切斷;alpha - 糖水解雙糖和寡糖裡的 1 → 4 糖鍵,結果,澱粉崩解的最終產物就是葡萄糖 (Radford et al.,1996)。
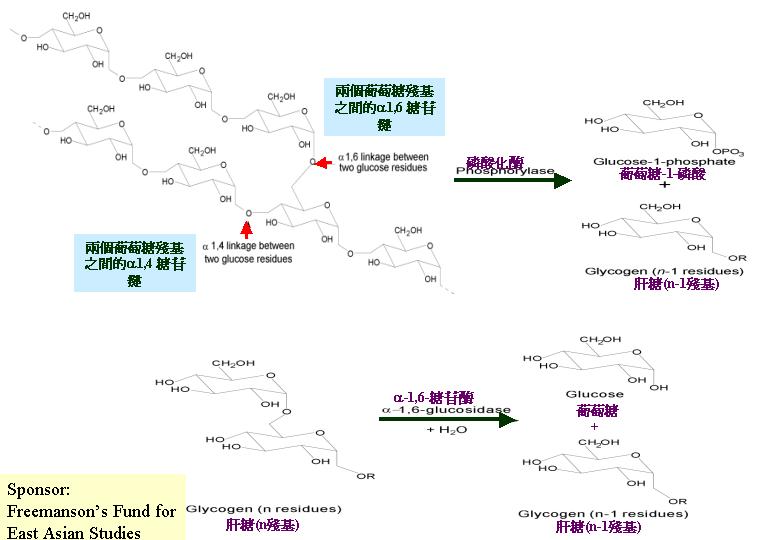
動物和真菌都以肝糖 (glycogen) 為多糖的儲備物,肝糖是葡萄糖以 alpha- 1 → 4 糖鍵聚合而成, 而每十個殘基便以alpha- 1 → 6 糖鍵分支開去。灰蓋鬼傘的子實層有詳盡的研究, McLaughlin (1974, 1977) 發現擔子下細胞先儲存多糖, 讓擔子分化之用,而在草菇 (Volvariella volvacea)
和鳳尾菇
(Pleurotus pulmonarius) 結實前和擔子果內,都有肝糖儲存 (Chiu
& Cheung, 1991; Chiu & To, 1993)。估計真菌會同樣利用澱粉系統去降解體外肝糖。至於體內肝糖就藉磷酸化作用,切斷 1→4 鏈,釋放葡萄糖 1-磷酸作新陳代謝之用,還有,轉移和alpha- 1 → 6 糖進行肝糖脫支 (breakdown
of glycogen)。真菌的肝糖以超微小粒結構在細胞質出現,粒上還可找到有關體。
除了肝糖外,有論說認為真菌利用細胞壁作儲存碳之用。
中間代謝 Intermediary metabolism, primary metabolism
真菌子實體營養菌絲提供各式各樣的代謝產物和生物量。其中藥用真菌具生物活性的
(bioactive) 謝物就包括:凝集素(lectins)
、多糖
(polysaccharides) 、多
polysaccharopeptides、等,其生物活性就非常多樣化,包括:降血糖、降血脂、抗癌、抗病毒、消炎、護肝等,其中靈芝、雲芝已廣受關注的
(Chiu et al., 2000)。
當所有的營養物是充裕時,真菌的生長速度會達到最高 (以 μmax表示),這時的代謝程序便是初生新陳代謝 (primary metabolism) (又名中間代謝; intermediary metabolism) 。次生代謝 (secondary metabolism) 是指真菌不在最佳生長狀態時而特有的代謝程序。所有新陳代謝的基本功能是轉化營養物為有用的生物能量 (ATP)、產生還原核甘酸輔 (NADH 和 NADPH) 和細胞組織所需物質(特別是大份子)的前體。
碳新陳代謝
Carbon Metabolism
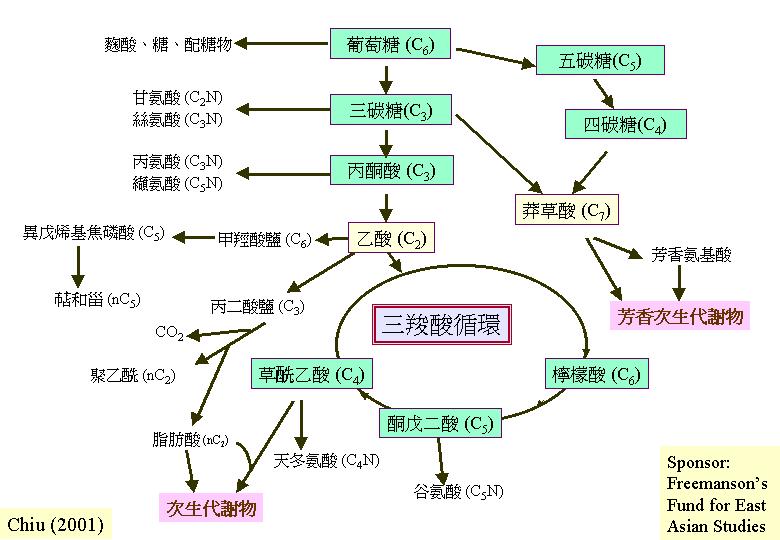
真菌和其他生物一樣:利用連串作用去進行碳新陳代謝,作用包括糖酵解 (glycolysis;HMP、EMP)、三羧酸循環 (tricarboxylic acid cycle,
Krebs cycle; TCA) 和氧化磷酸化作用 (oxidative phosphorylation)。葡萄糖的所有碳藉糖酵解作用和三羧酸循環以CO2釋放,而能量形式是三分子NADH2 、一分子FADH2和高能GTP,這些還原輔須經粒線體膜上電子傳遞體系恢復為ATP,這程序稱為氧化磷酸化作用。
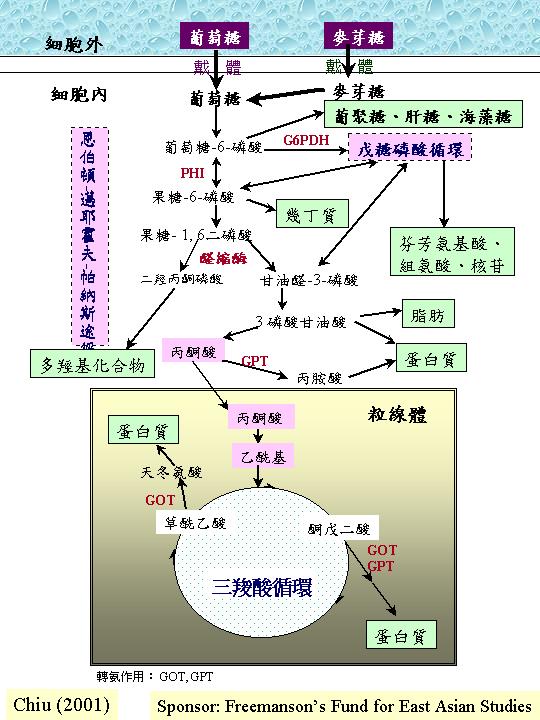
糖酵解作用:將葡萄糖轉變為丙酮酸 Glycolysis converts glucose to pyruvate
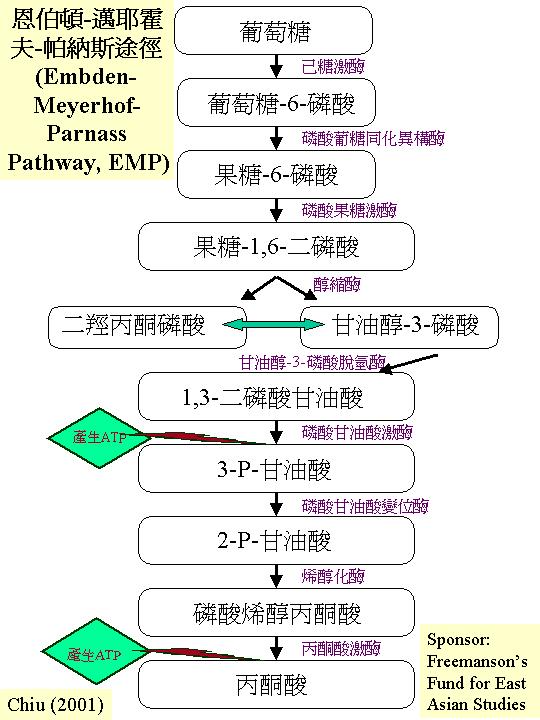
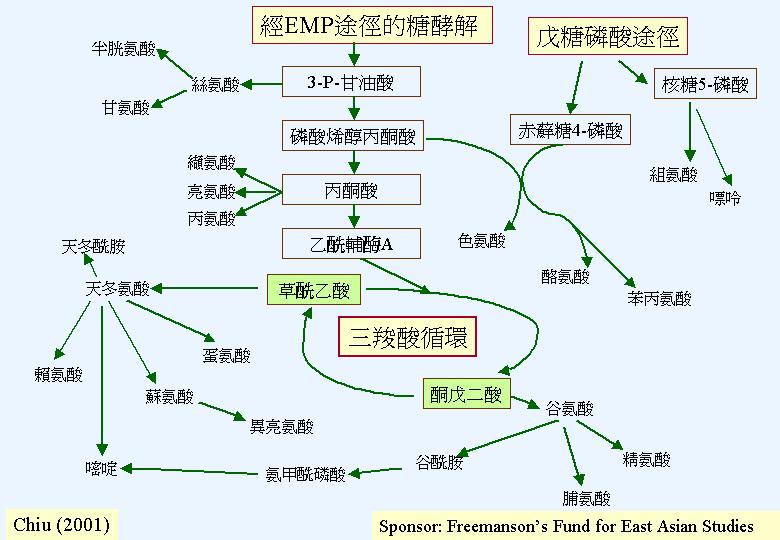
糖酵解作用只是描述葡萄糖轉變為丙酮酸,但絕不是逆轉的葡萄糖異生反應,其作用可有三路,其中之一為主路。大多數的真菌會採用恩伯頓-邁耶霍夫-帕納斯途徑 (Embden-Meyerhof-Parnass Pathway, EMP) ;這途徑的九個步驟都在細胞質進行。 結局是一分子葡萄糖轉變為二分子丙酮酸、二分子ATP和二分子NADH2。由此可見:恩伯頓-邁耶霍夫-帕納斯途徑產生的能量 (2ATP + 2NADH2)不高,反而它製造三羧酸循環所需的丙酮酸 (pyruvate)。而這路徑的標記 (marker enzymes) 是:磷酸葡糖同化異構 (phosphoglucose isomerase, PHI) 和醛縮 (
aldolase) 。
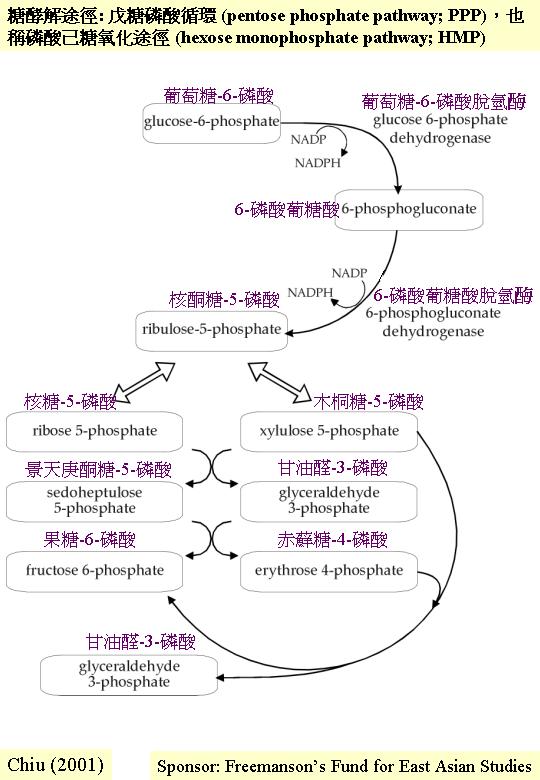
另一常見的糖酵解途徑是戊糖磷酸循環 (pentose phosphate pathway;
PPP),也稱磷酸己糖氧化途徑
(hexose monophosphate pathway; HMP)。 後稱謂更確切反映化學過程;葡萄糖-6-磷酸 (glucose 6-phosphate) 被轉化成果糖-6-磷酸 (fructose 6-phosphate) 和甘油醛-3-磷酸 (glyceraldehydes 3-phosphate),反饋進恩伯頓-邁耶霍夫-帕納斯途徑內。恩伯頓-邁耶霍夫-帕納斯途徑提供生產能量的主要中間產物 (丙酮酸) ,PPP則提供戊糖 (pentose sugar) 作核 (nucleotide) (包括輔和能量戴體、RNA和DNA) 合成之用,提供赤蘚糖磷酸 (erythrose phosphate) 藉莽草酸途徑 (shikimic acid pathway) 合成芬芳族氨基酸 (aromatic amino acids) 和提供NADPH2 (此輔是合成作用如脂肪和油的常用的還原劑)。 所以,雖然 PPP 能理論上達成完全的糖酵解 (六個周期就完全將葡萄糖氧化成CO2) , PPP 實容許多元化的生化網絡 (C metabolism)。而這路徑的標記 (marker enzymes) 是:葡萄糖-6-磷酸脫氫 (glucose-6-phosphate
dehydrogenase, G6PDH) 。
少數真菌使用恩特納-道德洛夫途徑 (Entner-Doudoroff (Edmund)
pathway, ED) 作糖酵解途徑,它經6-磷酸葡糖酸轉化成2 –酮基-3-去氧-6-磷酸葡糖酸
(2-keto-3-deoxy-6-phosphogluconate),最後產生丙酮酸 (pyruvate) 和磷酸甘油醛 (glyceraldehyde 3-phosphate) 。
細胞因應生長時間、階段、活動和營養所需的中間產物而作出調節,使用不同的糖酵解途徑作主導 (見Chapter
4)。大體上,快速生長的和分化的細胞都採用PPP,休眠和靜止細胞則減低PPP用率。在實驗中,以氨鹽基或硝酸作氮源會刺激細胞的 PPP 活動去生產更多的NADPH2。
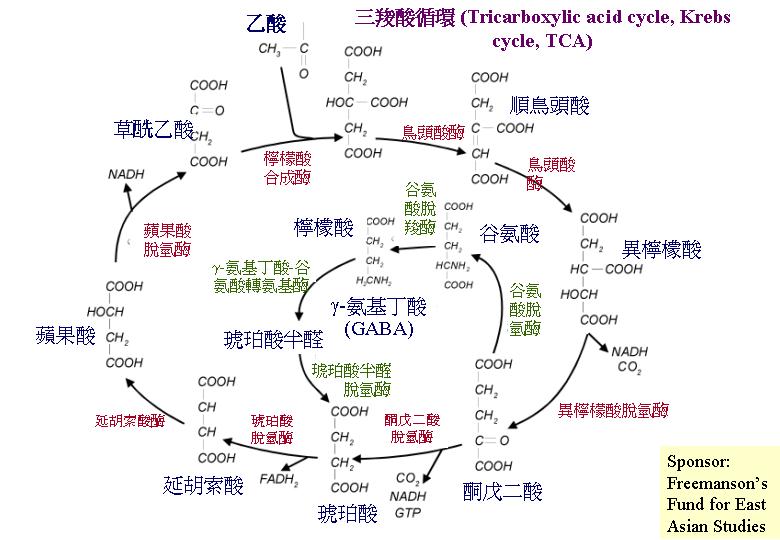
三羧酸循環 (Tricarboxylic acid cycle, Krebs cycle, TCA):丙酮酸的氧化
糖酵解作用在細胞質產生的丙酮酸會被傳送到線粒體,然後被轉換成乙輔A,負責的是:丙酮酸脫氫複合體 (先將丙酮酸脫羧,然後加上乙團體)。這反應序列是循環的,因為最後產物草乙酸會與乙輔A反應,將丙酮酸餘下的碳原子再推進三羧酸循環 (TCA) ,所以丙酮酸完全轉換成CO2,釋放 10質子和能量 (三分子NADH2 、一分子FADH2和高能GTP)。 三羧酸循環中的琥珀酸脫氫 (succinate dehydrogenase) 是在線粒體內膜的,而其他的位於線粒體間質。與其他生物不同是:谷氨酸脫羧環 (glutamate decarboxylation
loop) ,2–酮戊二酸 (2-oxoglutarate) 不被氧化脫羧成琥珀酸, 而是氨化成谷氨酸 (glutamate)。然後,谷氨酸脫羧成 4-氨基丁酸 (4-aminobutyrate);後者和2–酮戊二酸進行轉氨基作用 (transamination) 產生琥珀酸半醛 (succinate semialdehyde),這才被氧化成琥珀酸進入三羧酸循環上。灰蓋鬼傘 (Coprinus cinereus) 和雙孢蘑菇 (Agaricus bisporus) 的菌蓋就表現高活性的谷氨酸脫羧環 (Moore & Ewaze, 1976;
Moore, 1984a; Kumar & Punekar, 1997)。
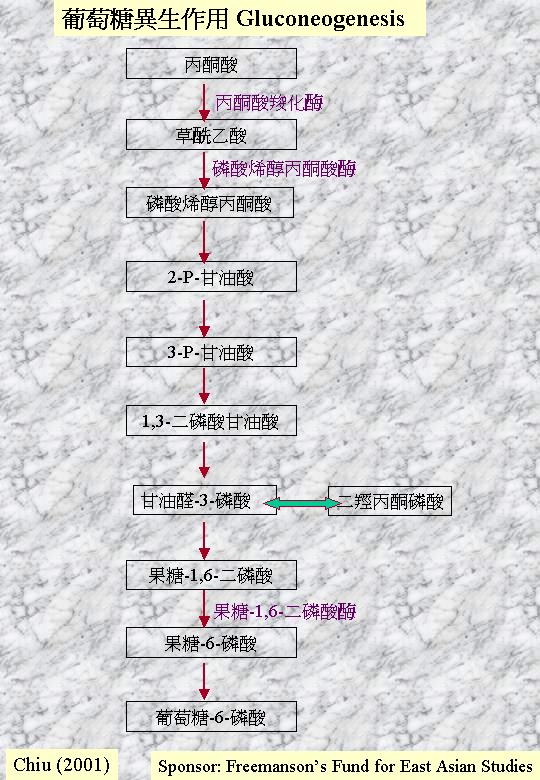
葡萄糖異生作用 Gluconeogenesis:合成碳水化合物
雖然透過糖酵解作用和三羧酸循環,真菌可利用多樣的碳和能量源,但如果生長在二粒碳原子的醋酸鹽 (acetate)上,真菌就要合成各樣碳水化合物,這不能靠逆向糖酵解作用,因為糖酵解作用的激 (kinases) 作用 (己糖激、磷酸果糖激和丙酮酸激)是不可逆轉的。 另一程式因而產生:葡萄糖異生作用;丙酮酸羧化 (pyruvate carboxylase) 將丙酮酸轉變成碳水化合物 (gluconeogenesis)。丙酮酸羧化將丙酮酸轉變成草乙酸 (oxaloacetate),然後磷酸烯醇丙酮酸羧化激 (phosphoenolpyruvate
carboxykinase) 將之脫羧和磷酸化成磷酸烯醇丙酮酸 (phosphoenolpyruvate),再經可逆的恩伯頓-邁耶霍夫-帕納斯途徑轉變成果糖-1,6-雙磷酸 (fructose bisphosphate),由果糖雙磷酸 (fructose bisphosphatase) 再變成果糖 6-磷酸。既然,糖磷酸是可互換的,這混合物便提供寡糖 (oligosaccharides)和多糖 (polysaccharides) 的合成。
糖酵解作用和葡糖異生作用明顯地是受嚴控去保持平衡的。磷酸果糖激 (phosphofructokinase) 是糖酵解作用的主要控制點 (EMP),而葡糖異生作用的果糖雙磷酸則受檸檬酸活化,但ATP則抑制雙磷酸。
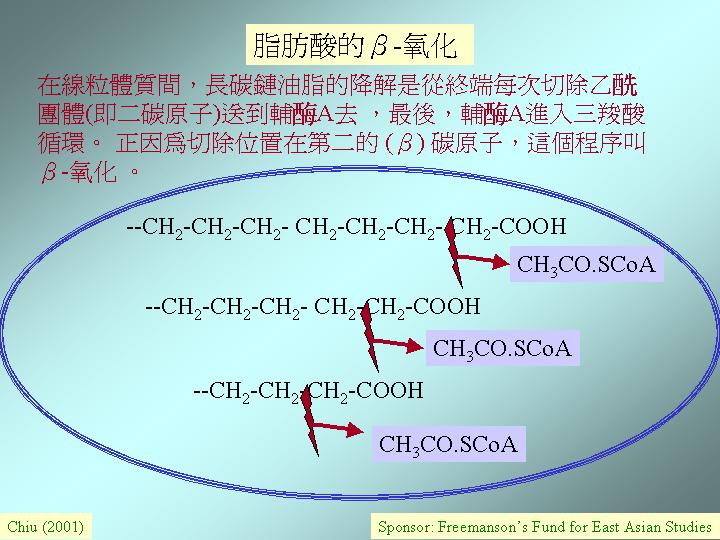
油脂的異化作用
油脂和肝糖都常用作儲存物,而油脂是非水溶的,並不影響滲透壓,所以是休眠期的孢子中常用的儲存形式。油脂被降解時,首步驟是脂肪將甘油的脂肪酸切除,後被轉成甘油醛-3-磷酸再送進糖酵解作用,脂肪酸只佔油脂分子量的 10%,其餘是長碳鏈 (如:棕櫚酸 (palmitic acid),C16;油脂酯酸 (stearic acid),C18)。這些鏈的降解是從終端每次切除乙團體(acetyl group, 即二碳原子)送到輔A(acetyl CoA) 去 ,地點在線粒體質間,最後,輔A進入三羧酸循環。 正因為切除位置在第二的 (β) 碳原子,這個程序叫β-氧化 (beta oxidation),同時,質子傳送到NAD 和FAD,例如:一分子棕櫚酸的降解需要七個切除點和產生八分子的乙-輔A,七個 NADH2 和七個 FADH2,後兩者進入氧化磷酸作用去釋放能量。結果,脂肪酸釋放大量能量;如:棕櫚酸的降解生產100分子的 ATP。因此,油脂是有效的能量儲存物。
氮新陳代謝
氮的吸收
真菌降解植物腐植質所吸收的主要氮源為蛋白質,其中包括:植物蛋白、木質素蛋白和微生物的蛋白。體外蛋白水解多底物,它將蛋白質和(縮氨酸)的鍵水解,切成細小的片斷直致氨基酸為止。蛋白質可作為碳、氮和硫源,當蛋白質作為唯一碳源時,培養瓶的胺基鹽 (ammonium) 濃度就會劇增,因為氨鹽基是由微生物使用蛋白質當碳源而排泄的產品。這些擔子真菌的體外蛋白主要受制於誘導作用;縱然環境可能有足夠氨、葡萄糖和硫酸的供應,只要蛋白質是可用的,真菌就生產蛋白,而生物量就有較高的增加( (Kalisz et al., 1986)。因此,蛋白質才是首選的底物。
合成氨基基團 (amino group)
生物的所有氮源本來自大氣的氮 (nitrogen)。每年固氮細菌和藍綠藻靠其固氮系統將100 至200噸的大氣氮轉為氨鹽基 (ammonium) 。如果真菌不能從環境直接吸收氨基酸的氨基基團,便會轉用氨鹽基。這氨鹽基同化過程便得依靠真菌普遍存在的谷氨酸脫氫 (glutamate dehydrogenase,
GDH),它催化氨鹽基和2-酮戊二酸合成谷氨酸。真菌的谷氨酸脫氫有兩類:一連結到NAD和另一個連結到 NADP。
氮在細胞內會被重新分配 (N metabolism) ,2–酮戊二酸 (2-oxo-glutarate) 和谷氨酸 (glutamate) 的互相轉換是細胞新陳代謝的中心,而且也是碳新陳代謝和氮新陳代謝的交枝點。這反應是可逆向的;NAD - 連結谷氨酸脫氫 (NAD-GDH) 有脫氨基或分解代謝功能 (谷氨酸 → 2–酮戊二酸 + 氨鹽基),相反,NADP-連結谷氨酸脫氫提供胺化或合成代謝的功能 (2–酮戊二酸 + 氨鹽基 → 谷氨酸)。
的路徑,顯示氨基酸、嘌呤和密定的氮源。
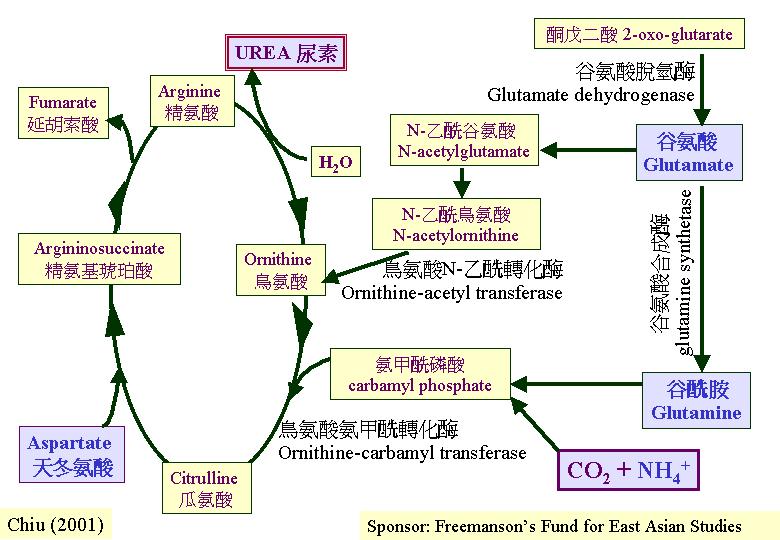
處理過剩的氮
活細胞內各種物質都在持續不斷地流動和被周轉;舊的會被分解代謝,而新的會被合成。餘碳會以CO2形式排放,餘氫則以水形式,而餘氮當氨鹽基 (ammonium) 或尿素 (urea) 形式。陸生的哺乳動物就將代謝蛋白質而產生的氨鹽基,利用尿素周期產生尿素去排泄和逃避氨鹽基的毒性 (N disposal) 。真菌菌絲期的尿似是一組成,所以,任何被形成的尿素會分解成氨( NH3 )和 CO2。只有少數情況下,尿素會出現積聚如在擔子果中 (Reinbothe & Tschiersch,
1962; Reinbothe et al., 1967;Moore, 1998; Chiu & Moore,
1999) ,尿素這時的功能是控制細胞的滲透壓。
尿素循環即以尿素排除餘氮的過程 (N disposal),灰蓋鬼傘 (Coprinus cinereus) 發育中的菌蓋表現活躍的尿素循環代謝和積聚尿素。尿素循環由四個去阻遏的活動組成,這四個就是: NADP-谷胺酸脫氫(NADP-GDH)、谷胺酸鹽合成 (glutamine synthase, GS)、鳥胺酸乙轉移 (ornithine-acetyl transferase, OAT)和鳥胺酸氨基甲轉移 (ornithine-carbamyl transferase, OCT)。合成氨甲磷酸(carbamyl phosphate)的氨鹽基分子和天冬氨酸 (aspartate) 的氨基最後被「棄」進尿素分子內。尿素吸引水的滲透,水進入菌蓋引起細胞膨脹,導致傘蓋擴張,這便是灰蓋鬼傘和鳳尾菇菌蓋主要的形態變化 (Moore, 1998; Chiu & Moore, 1999)。
次生代謝 Secondary Metabolism
次生新陳代謝產物這名詞可追溯到J. D. Bu’Lock (1961) ,他將新陳代謝分為初生和次生新陳代謝: 初生的所有生物都有同一代謝模式,次生的分佈較狹 (可能只限某一品種) 而不一定有明顯功能。對次生新陳代謝認知的困難,原因有:
(i)
我們對這代謝的生化和生理都沒有資料, [或]
(ii)
大部份有經濟效益的次生代謝產物是透過人工栽培而非自然環境下產生, 所以不能知悉它的天然功能。
次生新陳代謝的通常特徵是:它由連串小的反應連接而成一程序,將初生代謝的一些中間產物轉換成各式各樣的產品 (Bu’Lock, 1967) (metabolites),即次生代謝較後階段的反應是獨特的 (Campbell, 1984)。
初級代謝和次生代謝的變化關係:初次和次生代謝可同時在相同的細胞發生而且利用相同的碳中間物, 然而後在許多方面都表現複雜的生物化學。但是多數的次生代謝都利用化學途徑上各小步驟的中間產物;乙輔A(acetyl CoA) 也許是次生新陳代謝最重要的底物,它可轉成松烯 (terpenes)、類固醇 (steroids)、脂肪酸 (fatty acids) 和多烯酮 (polyketides)。磷酸烯醇丙酮酸和赤蘚糖-4-磷酸藉莽草酸途徑合成芳香族氨基酸和其他芳香產物,還有,其他的次生代謝物源於非芳香族的氨基酸 (metabolites)。
次生代謝物常指以下的途徑及其產物:
(i)
麴酸 (kojic acid)、糖、配糖類 (glycosides) belonging to
glucose-derived metabolites
(ii)
丙二酸途徑melonate pathway
(iii)
甲羥戊酸途徑mevalonic acid pathway
(iv)
莽草酸途徑shikimic acid pathway
(v)
芳香族氨基酸和其他芳香產物/氨基酸衍生物 (amino acid-derived
metabolites /aromatic secondary metabolites) 。
在醱酵工業中,液體醱酵酵所生產的產品非常繁多,如:釀酒、製、維他命、抗生素、降血脂劑等等,最近,更以 air-lift fermentation
technology 氣升醱酵技術生產「真菌肉」(mycomeat) 作素食食物 (vegetarian food) (Moore
& Chiu, 2001) 。而固體醱酵的應用,主要是化廢為寶,將世界最大量的植物纖維廢料轉化成可食用或藥用真菌,如擔子果等。因此,真菌的生化是重要的研究和提升的對象。既然營養是從外界吸收和提供的,這便成為操控真菌生化的手段,真菌亦成為醱酵工業的重要成員。
甲羥戊酸途徑mevalonic acid pathway: 合成類、類胡蘿卜素和固醇
靈芝 (Ganoderma lucidum, Ganoderma
tsugae, Ganoderma sinense) 的抗癌、抗衰老代謝物多是類。類 (terpenes)、類胡蘿卜素 (carotenoids) 和類固醇 (steroids) 都有一五碳構造如異戊二烯 (isoprene)。
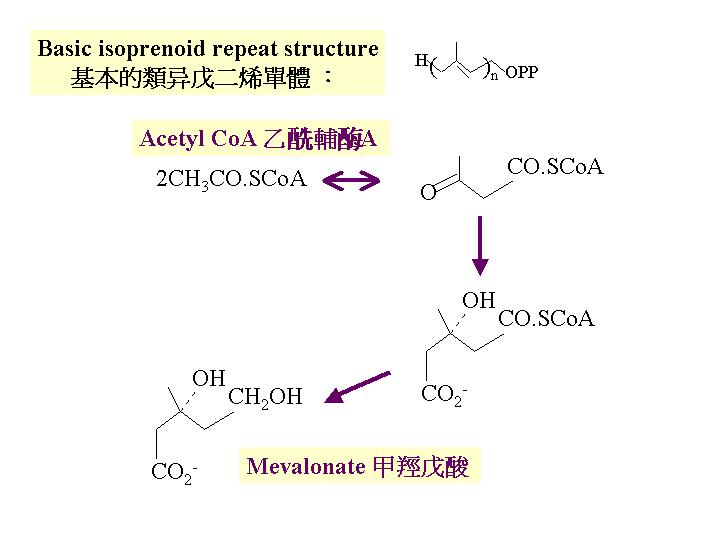
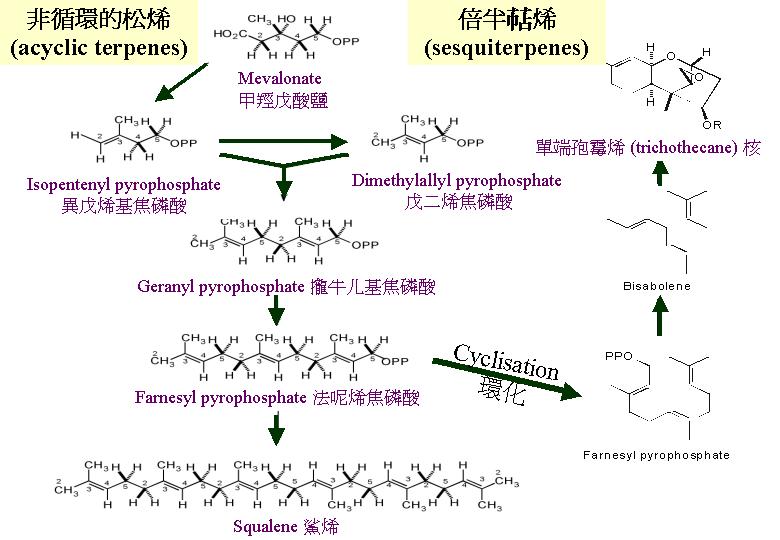
二分子的乙輔A合成甲羥戊酸 (mevalonate)。類 (terpenes) 是異戊二烯 (isoprene) 單位的衍生物,類是最簡單的天然類異戊二烯混合物,它透過將異戊二烯單體縮合而連接成鏈狀 。的狹義專指C9H16 化學物, 但廣義是指: 含十個異戊間二烯以上的化學物,可以是鏈狀或環狀的。鏈狀由異戊二烯二聚體與類戊焦磷酸縮合而成一9碳的單焦磷酸牻牛几酯 (terpenes)。真菌最大類的是和倍半多烯(sesquiterpenes) ,為一類環狀的。 它的碳骨架可能是靠焦磷酸法呢酯環化時產生。
經連串的縮合作用合成非循環的松烯 (acyclic terpenes),甲羥戊酸可轉化成戊二烯基焦磷酸 (dimethylallylpyrophosphate)
和異戊基焦磷酸 (isopentenylpyrophosphate) ,後兩者的縮合形成攏牛儿基焦磷酸 (geranyl pyrophosphate) ,再合成法呢烯焦磷酸。而法呢烯焦磷酸的環化可能製造單端孢霉烯 (trichothecane) 核,合成倍半烯 (sesquiterpenes)。
而雙 (diterpenes) 則靠焦磷酸牻牛几基牻牛几酯環化而成,雙烯攏牛儿醇攏牛儿基焦磷酸的環化產生赤烷 (gibbane) 和松烷 (kaurane) 的結構。 這類產品包括植物病害真菌近粘藤倉赤霉 (Gibberella fuijikuroi)
[串珠鐮孢
(Fusarium moniliforme)的有性階段] 產生的一植物激素:赤霉素 (gibberellic acid)。
三 (C30)的重要性是其鏈狀成員鯊烯 (squalene) 是固醇 (甾醇,steroids) 的前體。固醇(和環狀三) 就源於氧化鯊烯環化縮合而成。麥角固醇 (ergosterol) 是真菌獨有, 它便可成為抗真菌劑的抑制目標 (Barrettbee & Dixon,
1995)。
Cytochrome
P450轉化多類化學物,包括:合成體內的固醇 (steroids) ,如:真菌獨有的麥角固醇 (ergosterol) 等,其中主要的 (lanosterol 14α-demethylase) 是屬於細胞色素P450家族cytochrome P450 superfamily的。
與鯊烯相似,C40的類胡蘿卜素是由8個異戊二烯單位 (四類)以頭尾相連方式縮合組成。這類色素包括:非環狀的番茄紅素,單環、雙環的γ-胡蘿卜素和 β-胡蘿卜素。類胡蘿卜素和一些酮基衍生物在真菌界廣泛但不是絕對存在 (Goodwin, 1976; Armstrong
& Hearst, 1996) ,因而可應用為分類的標準 (Valadon, 1976)。
丙二酸途徑 (malonate pathway, polyketide pathway) :合成聚乙及其衍生物
大部份的真菌次生新陳代謝產物是由丙二酸途徑合成的 (Turner, 1976)。聚乙 (polyketides) 由多個酮甲基縮合而成的非環狀鏈組合而成 (poly-beta-ketomethylenes)。如類一樣可摺疊和環化產生廣泛不同的大分子 。聚乙單體的乙基團 (acetyl group, -CH2CO- unit) 由乙輔A提供,所以,聚乙的合成與脂肪酸的有許多相同處。
聚乙及其衍生物的鏈能以多種方式摺疊,若和內在的醇醛 (aldol) 縮合,形成密閉芬芳環的結構。丁烯酮 (tetraketide) 的各種環化,可產生:苔色酸 (orcellinic acid) 和乙間苯三酚 (acetylphloroglucinol) 。而戊烯酮 (pentaketide) 的五種環化方式,結果是:煙曲霉 (fumigatin) 、盤尼西林酸 (penicillic acid) 、棒曲霉素(patulin) 、stipitatic acid 和毒素橘霉素 (citrinin) 。多聚乙 (polyketides) 的例子有: LL-Z1272a 是鐮刀菌 (Fusarium) 產生的抗生素,其聚乙衍生芬芳環有一倍半烯的取代基,黃曲霉素 (aflatoxin B1) 、格鏈孢酚 (alternariol) 等。
有趣的是:紅米 (Monascus) 的色素 (pigments) 和毒素橘霉素(citrinin)都是聚乙產物 (Hajjaj et al., 1999) ,但前者是傳統食物添加劑,後者是有損腎、肝功能的毒素,因此代謝調控是重要的。另外,許多屬如: Aspergillus, Pencillium
和Monascus都生產橘霉素,但只有Monascus以丁烯酮 (tetraketide) 作橘霉素(citrinin) 的前身,Monascus以戊烯酮 (pentaketide) 為中間產物去製造色素 (Hajjaj et al., 1999)。可見另類的代謝多樣性 (metabolic diversity) 。
其他次生新陳代謝途徑
莽草酸-分支酸途徑 (shikimate-chorismate
pathway) 合成多種芬芳化合物和一些芳香族氨基酸,包括: 苯丙氨酸 (phenylalanine)、色氨酸 (tyrosine)、和酪氨酸 (tryptophan) (metabolites),同時也提供其他芳香族混合物的中間產物 (Turner,1971; Towers, 1976)。 許多真菌和植物製造簡單的代謝物,如:五倍子酸(gallic acid) 、 焦酚 (pyrogallol) 和p-甲氧基肉桂酸 (p-methoxycinnamate) 。
重要的色氨酸衍生物有:麥角菌 (Claviceps purpurea)
生產的麥角亭
(ergocristine) 屬於麥角症生物鹼、致幻劑: 麥角二乙基胺和摩術蘑菇 (Psilocybe) 的裸蓋菇素 (psilocybin) 等。 而圓孔中肝菌 (Gyroporus cyanescens)
在子實體受傷而出現的藍色色素是(gyrocyanin) 的氧化物,硬毛多孔菌 (Polyporus hispidus) 由硬毛素 (hispidin) 聚合物使子實體變堅固。
非芬芳氨基酸類衍生的次級新陳代謝物,包括: 毒蕈 (Amanita muscaria) 的主要毒素:毒蠅素 (muscarine) 和白僵菌素 (muscaridine),它們是從谷氨酸 (glutamic acid) 合成的,蘑菇屬的蘑菇毒素 (agaritine) 和谷氨胺羥色苯 [glutaminyl hydroxybenzene,簡稱 GHB;與 p-hydroxy(γ-glutamyl) anilide同名] 是 N-化 (N-acylated) 的谷氨酸類分子。蘑菇毒素可佔其子實體乾重的0.3%,而 GHB 可能控制擔孢子的休眠和為擔孢子細胞壁色素的前體 (Hammond & Wood, 1985)。擬青霉 (Paecilomyces
varioti) 所產生的擬青霉素 (variotin) 有抗真菌活性,屬γ-氨基丁酸 (γ-aminobutyric acid) 與六烯酮 (hexaketide) 的N-化衍生物。至於N-雙甲基甲硫氨醇 (N-dimethylmethioninol) 是沙門柏干酪青黴菌 (Penicillium
camemberti) 將甲硫氨酸 (methionine) 脫羧後形成的揮發性胺 (amine),它是法國沙門柏干Camembert 芝士 (軟質乳酪) 的濃香物質。
另外,盤尼西林類抗生素代表縮氨酸 (peptides)衍生的次生新陳代謝產物。由盤尼西林的前體:δ-(α- 氨-脂肪)半胱氨纈氨酸,和纈氨酸殘基結合,使衍生盤尼西林核。
脂肪酸的修飾可生產如:聚乙炔 (polyacetylenes) 和 環戊烷 (cyclopentanes) 等多種次級代謝物。 當乳菇 (Lactarius) 子實體受傷時,流出的乳液就有乳菇酸 (lactarinic acid),乳菇酸是一6酮硬脂酸 (ketostearic acid),它和真正的橡膠基質(異戊二烯聚合物)是不同的。
營養消耗是最基本誘發次生新陳代謝的,各式營養物就引起不同的途徑。Bu’Lock (1961) 提議:細胞繁殖籍提供一個不斷擴張初生新陳代謝的儲池去推動平衡生長,但當生長停止、特別是維持生長期時,次生新陳代謝便提供另類儲池:好讓合成和營養吸收的機制繼續下去,縱然其產物沒甚功能。但是,次生代謝物有其品種的專一性,這點不能以或然率去解釋。Campbell (1984) 提出類似(並不排外的)論點:既然次生新陳代謝是那麼複雜,它定有進化意義。達爾文認為同一屬中各品種和同一品種的各亞種中的競爭是極其激烈的,於是,這類分佈狹窄的代謝物就給予品種或亞種優勢。
動物少有次生新陳代謝,但動物也生產毒素、抗毒素、激素和類激素等,這些動物天然物質不被稱次生代謝物 (secondary metabolites),因為我們在未有提純化學物時,我們已知道它們的功能。其實,這只是詞匯的問題;其一例子便是:蜘蛛分泌一幼蟲類激素去吸引獵物,這現象動物學家稱為化學擬態,實與次生代謝無異 (Stowe et al., 1987) 。另一例子是:蟾蜍 (Bufo)分泌的、引起幻覺的蟾蜍毒素 (bufotenin),毒蕈捕蠅蕈屬 (Amanita) 也生產 (Metzenberg, 1991),那麼,為何同一代謝物在不同的個體有不同的稱謂?
鳴謝:
承蒙遠東及香港區共濟會之菲裘新研究東亞學科基金的贊助,使這部介紹真菌生理、生化、遺傳、細胞學、發育和形態及生態的資料可以中文出版,著者深致謝意! 特別鳴謝W. Bro. Peter J. Nunn 先生的支持。
The authors are deeply grateful to The Freemasons’ Fund for East Asian Studies by the District Grand Lodge of Hong Kong and the Far East, without which this publication would not have been possible. We are especially grateful to Mr. W. Bro. Peter J. Nunn, District Grand Secretary of the Fund for the kind support.
參考書目 Reference
Barrasa, J. M., Gutiérrez, A., Escaso, V.,
Guillén, F., Martínez, M. J. & Martínez, A. T. (1998) Electron and
fluorescence microscopy of extracellular glucan and aryl-alcohol oxidase during
wheat straw degradation by Pleurotus eryngii. Applied and Environmental
Microbiology 64, 325-332.
De Jong, E., Field, J. A., Spinnler, H.E.,
Wijnberg, J. B. P. A. & de Bont, J. A. M. (1994) Significant biogenesis of
chlorinated aromatics by fungi in natural environments. Applied and
Environmental Microbiology 60, 264-270.
Jeffers, M. R., McRoberts, W. C. & Harper, D.
B. (1997) Identification of a phenol- 3-O-methyltransferase in the
lignin-degrading fungus Phanerochaete chrysosporium. Microbiology
143, 1975-1981.
Leonowicz, A., Matuszewska, A., Luterek, J., Ziegenhagen,
D., Wojtaś-Wasilewska, M., Cho, N.-S., Hofrichter, M. & Rogalski, J. (1999)
Biodegradation of lignin by white rot fungi. Fungal Genetics and Biology
27, 175-185.
Moore, D. (1998) Fungal Morphogenesis.
Cambridge University Press.
Palma, C., Martinez, A. T., Lema, J. M. &
Martinez, M. J. (2000) Different fungal manganese-oxidizing peroxidases: a
comparison between Bjerkandera sp. and Phanerochaete chrysosporium.
Journal of Biotechnology 77, 235-245.
Zacchi, L., Morris, I. & Harvey, P. J. (2000)
Disordered ultrastructure in lignin-peroxidase-secreting hyphae of the
white-rot fungus Phanerochaete chrysosporium. Microbiology 146,
759-765.
If
you use diagrams or material from this website, please cite the home page and
authors or the appropriate source publication in your work.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}