
3.8 Basidiomycota
Basidiomycota (informally, basidiomycetes) is another large group, comprising about 32,000 known species. Morphologically, ecologically, and taxonomically this is a very diverse group, but its members share the feature that their sexual spores are exospores, formed on a basidium (and are therefore called basidiospores).
Basidiomycota includes the plant pathogens that cause smut and rust diseases, ectomycorrhizal species which are of key significance to forest ecosystems, saprotrophic species that can decay the lignin (‘white-rot fungi’) as well the cellulose of plant litter, and the most noticeable and frequently encountered mushroom fungi. Several basidiomycetes have mutualistic associations with insects (e.g. leaf-cutter ants, termites, ambrosia beetles) that are based on the ability of the fungus to digest plant litter efficiently.
All the fungi whose fruit bodies are commercially farmed as food are basidiomycetes (truffle ‘cultivation’ is different, being more a matter of the silviculture of the truffle groves; see Section 11.6). On the other hand, some members of this group produce toxins that can be hallucinogenic (e.g. Psilocybe cubensis) or deadly poisonous (e.g., the ‘Destroying Angel’, Amanita virosa). The species that causes cryptococcal meningitis in persons suffering compromised immunity (due to HIV infection, cancer chemotherapy, metabolic immunosuppression to maintain a transplanted organ) is also a basidiomycete; the disease is caused by the asexual form (called the anamorph), which grows as a yeast. This form is given the generic name Cryptococcus. The sexual form or teleomorph is called Filobasidiella.
The Basidiomycota divides into three distinct evolutionary groups which are given Subphylum status in the AFTOL study (Blackwell et al., 2006; Hibbett et al., 2007) and this still represents the most widely accepted classification scheme (and see He et al., 2019 and Varga et al., 2019) for the most recent, and extensive, phylogenetic treatments):
- Ustilaginomycotina (smuts and allies; the traditional ustilaginomycetes).
- Pucciniomycotina (rusts and allies; the traditional urediniomycetes).
- Agaricomycotina (the traditional hymenomycetes or basidiomycetes and allies: producing a macroscopic fruit body with a hymenium spread over gills (as in mushrooms), within pores (boletes and bracket fungi), or over a toothed (hedge hog fungi), coralloid (coral fungi), labyrinthoid (daedaleoid fungi), wrinkled (merulioid fungi), or smooth to diffusely encrusted (corticioid fungi = resupinate fruit bodies) tissue structure; or completely enclosed (gasteroid fungi).
- To these may be added the class Wallemiomycetes (incertae sedis, which is Latin for ‘of uncertain placement’) which comprises three species of fungi in the genus Wallemia and is a 500-million-year-old sister group of Agaricomycotina (Padamsee et al., 2012; Zajc et al., 2013). These are the most halophilic fungi known; Wallemia ichthyophaga requires at least 1.5 M NaCl for growth in vitro and thrives even in saturated NaCl solution. A limited number of strains of Wallemia have been isolated so far; from salt pans used to crystallise salt from sea water (‘solar salterns’) and salted meat (Zajc et al., 2016).
Ustilaginomycotina is a group of about 1,200 fungi in approximately 62 genera, most of which parasitise plants, mainly non-woody angiosperm herbs, especially grasses (Poaceae) and sedges (Cyperaceae). The best known members, Ustilago and Tilletia, cause diseases of cereals known as smut or stinking smut that can severely reduce crop yield. Ustilago maydis, which causes smut disease on maize (Zea mays), is used widely as a model organism for plant pathogenesis and was the first basidiomycete plant pathogen to have its complete genome worked out (Begerow et al., 2014).
In these fungi the basidiospores establish a yeast-like budding haploid phase (the cells are called sporidia) that grows saprotrophically (Fig. 4); eventually conjugation of compatible haploid cells produces a dikaryotic, parasitic mycelium that infects the host plant.
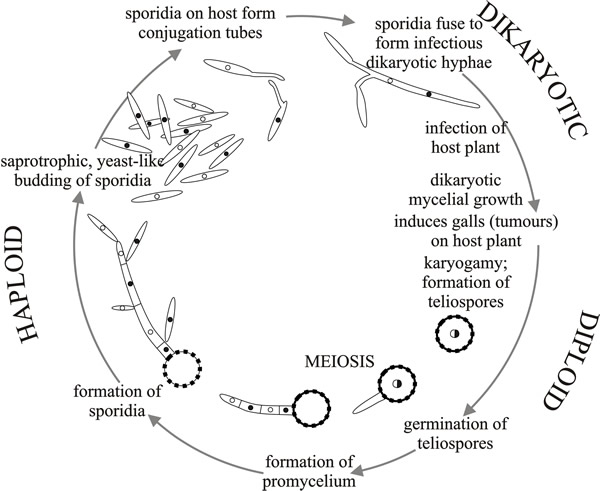 |
| Fig. 4. Diagram of the life cycle of Ustilago maydis. From Moore & Novak Frazer, 2002. |
Mating (controlled by mating type factors, see Section 8.5) is essential for infection of host plants, and the dikaryotic plant-parasitic phase ends with the production of teliospores, which, in most of these fungi, develop thick-walls and are the dispersal spore.
Teliospores are formed in a sorus (a sorus is a mass of spores that bursts through the host epidermis; the word comes from the Greek soros, heap) which is on or in the parenchyma tissue of the host. The location depends on species; sori may appear on roots, stems, leaves, flowers, seeds, etc. As the teliospores are usually powdery and dark brown or black, the plant looks as though it is covered in particles of dirt or soot, so giving rise to the common name ‘smut disease’.
Other names to look out for are Graphiola, Exobasidium, and Microstroma. The group also includes some animal pathogens in the genus Malassezia, which are lipophilic yeasts (no dikaryophase known) isolated from the skin of warm-blooded animals. These fungi require fat to grow and are found in areas of the skin with many sebaceous glands: scalp, face, and upper part of the body. In humans, Malassezia globosa causes dandruff and seborrhoeic dermatitis. It is possible that Malassezia species originated from a dikaryophase parasitic on plants.
Roughly 8,400 of the species assigned to Basidiomycota belong to Subphylum Pucciniomycotina. The Pucciniomycotina differs from the other two Subphyla by the possession of simple septal pores lacking membrane-bound caps, and by the sugar composition of the cell wall. Most members of the Pucciniomycotina are parasitic, and about 90% belong to a single Order of plant parasites, Pucciniales, known as rust fungi, that cause some of the most devastating diseases of crops; but the group also includes pathogens that attack other fungi and those that attack insects (Aime et al., 2014). CLICK HERE to see a page of illustrations.
The rusts form a natural (monophyletic) group. Names to look out for include Puccinia (P. graminis causes stem rust of cereals), Uromyces (U. appendiculatus causes a rust disease in both temperate and tropical crops of the common bean, Phaseolus vulgaris). Endophyllum has only been recorded as a rust disease of the perennial woody shrub Chrysanthemoides monilifera; both host and rust originate in South Africa, but Chrysanthemoides is an aggressive weed in southern Australia and Endophyllum is showing promise as a biocontrol agent.
Basidiomycetous yeasts include Rhodosporidium (and its anamorph Rhodotorula), and Sporidiobolus (and its anamorph Sporobolomyces), which can be commonly isolated from surfaces in the domestic environment.
The Agaricomycotina (traditionally known as hymenomycetes or basidiomycetes) is a diverse group that includes about 21,400 described species, which is about 65% of known Basidiomycota (or about a fifth of all fungi). The Agaricomycotina probably originated between 380 and 960 million years ago. Today the group includes many wood decayers, litter decomposers and ectomycorrhizal fungi. It also includes a lesser number of important plant pathogens, one of which is the largest organism on the planet. The AFTOL classification scheme for the Subphylum Agaricomycotina is listed below with hyperlinks to illustrations. Here we will briefly describe the major groups.
The largest clade is the Order Agaricales or euagarics clade (in the Subclass Agaricomycetidae), which is made up of the mushroom forming fungi and includes more than half of all known species of the homobasidiomycetes, like the type genus Agaricus (Kerrigan, 2016). More than 13,200 species; and roughly 410 genera have been ascribed to the 33 families of the order. Traditionally, the shape and morphology of the mature fruit body, the colour of the spore deposit, and various anatomical and cytological features have figured largely in the taxonomy; but because apparently similar structures and shapes can evolve in different ways this approach led to the establishment of many artificial groups. For example, there are several ways of creating the folded surface that we call gills. Since they arise in different ways such features are only superficially similar because they have evolved in different ways (like, for example, the wings of butterflies and wings of birds); they are described as analogous organs (as distinct from homologous organs, which are those that have a shared evolutionary ancestry). Shape and morphology of mature fruit bodies, even though these are the easiest to find in the field, can be misleading because they do not contribute to a natural classification. This is why introduction of molecular sequence comparisons is prompting major, and sometimes surprising, revisions in classification of these organisms. Phylogenies based on analysis of ribosomal RNA sequences has transformed the classification of the Agaricales, and interestingly, has shown that ecological traits have been underused in the past in diagnosing natural groups [CLICK HERE to see illustrations of Agaricales].
Another major group of mushroom-forming fungi that has representatives in most forest ecosystems around the world is the Order Boletales. This order contains around 1,320 described species although, as it is most diverse in the still under-researched tropics, this probably underestimates the real biodiversity. The Boletales includes conspicuous stipitate-pileate forms (‘mushrooms’ with stems and caps) that mainly have tubular, but sometimes lamellate (gilled) hymenophores, as well as intermediates that show transitions between the two types of hymenophore structures. The Boletales also includes puffball-like forms (that used to be called gasteromycetes), resupinate or crust-like fungi that produce smooth, merulioid (wrinkled to warted), or hydnoid (toothed) hymenophores. As well as diverse morphologies, the Boletales exploit diverse habitats, including wood decay but they differ from their sister clades (Agaricales and Atheliales) by not having white-rot fungi in the group. Instead, saprotrophic Boletales have developed a unique brown-rot that is especially adapted to decaying conifer timber. The majority of Boletales form mycorrhizal associations and some are mycoparasites [CLICK HERE to see illustrations].
Some of the most eccentric fruit body morphologies occur in the Hysterangiales, Geastrales, Gomphales and Phallales (combined in a grouping known as gomphoid-phalloid fungi (meaning ‘like Gomphus and/or like Phallus’; CLICK HERE to see illustrations). This is where we find the cage fungi, the stinkhorns and puffballs (Fig. 3.18). The Gomphales contains both gasteroid (fruit bodies ‘stomach-like’ with completely enclosed hymenium) and nongasteroid taxa, with gasteroid morphology being derived from epigeous taxa (epigeous means occurring above the surface of the ground; contrasts with hypogeous, which describes underground or semi-underground fruit bodies). On the other hand, in the Phallales the truffle-like hypogeous form is an ancestral morphology of the stinkhorn fruiting bodies. Although basidiospore maturation occurs within the enclosed young primordial fruiting bodies of the stinkhorn (the ‘eggs’), lifting the mature spore producing tissue into the air where flies can easily find it represents an independent instance of the origin of stipes within the Basidiomycota.
The Cantharellales, which includes mushroom fungi like Cantharellus and Craterellus [CLICK HERE to see illustrations] was set apart from other gilled fungi very early in the history of mycology on the grounds that the gills are formed when the hymenophore folds into pleats like a fan, so these were called ‘false gills’ in comparison with the allegedly ‘true gills’ of most other mushrooms (understood to be individual, plate-like or blade-like things, structurally separate from one another and from the flesh of the cap). In today’s phylogeny the difference is still important, but is less surprising because we recognise the lamellate (gilled) hymenophore as a simple strategy to increase the area available for spore formation, and, more importantly, it’s a strategy that can be arrived at through several different evolutionary routes.
The Hymenochaetales, which is dominated by wood-decaying species, also includes many variations of fruit body types. Most species have basidiomata that are effused (stretched out flat) or effused-reflexed (with the edge turned up or turned back); a few form stipitate mushroom-like (agaricoid), coral-like (clavarioid) and spathulate (shaped like a spoon) to rosette-like basidiomata [CLICK HERE to see illustrations].
Most species have vegetative (sterile) cells in the fruit body tissue, often accompanying the basidia in the hymenium. Although these could collectively be called cystidia, many unique terms have been introduced for them to describe their distinctive morphologies; a simple example is that most species in the family Hymenochaetaceae have characteristic cystidia called setae. Because microscopic distinctions like these, and fruit body shapes, formed such an important basis for the traditional classification of fungi, what is now known as the hymenochaetoid clade draws its members from several traditional families: Agaricaceae, Polyporaceae, Corticiaceae, Stereaceae and Hymenochaetaceae, but includes only the type genus for the last family mentioned.
The Russulales is, probably, the most morphologically diverse group, because it contains resupinate, discoid, effused-reflexed, clavarioid, pileate, and gasteroid fruit bodies, and hymenophores that may be smooth, poroid, hydnoid, lamellate, or labyrinthoid. Members of the Russulales are primarily saprotrophs but others are ectomycorrhizal, root parasites and insect symbionts [CLICK HERE to see illustrations].
The AFTOL classification scheme for the Subphylum Agaricomycotina looks like this:
- Agaricomycetes
- Agaricomycetidae
- Agaricales [CLICK HERE to see illustrations]
- Atheliales [CLICK HERE to see illustrations]
- Boletales [CLICK HERE to see illustrations]
- Phallomycetidae
- Geastrales [CLICK HERE to see illustrations]
- Gomphales [CLICK HERE to see illustrations]
- Hysterangiales [CLICK HERE to see illustrations]
- Phallales [CLICK HERE to see illustrations]
- Agaricomycetidae
- Dacrymycetes
- Dacrymycetales [CLICK HERE to see illustrations]
- Tremellomycetes
- Cystofilobasidiales
- Filobasidiales
- Tremellales [CLICK HERE to see illustrations]
- Agaricomycetes [incertae sedis (= position still
uncertain)]
- Auriculariales [CLICK HERE to see illustrations]
- Cantharellales [CLICK HERE to see illustrations]
- Corticiales
- Gloeophyllales [CLICK HERE to see illustrations]
- Hymenochaetales [CLICK HERE to see illustrations]
- Polyporales [CLICK HERE to see illustrations]
- Russulales [CLICK HERE to see illustrations]
- Sebacinales [CLICK HERE to see illustrations]
- Thelephorales [CLICK HERE to see illustrations]
- Trechisporales [CLICK HERE to see illustrations]
Names to look out for include Phellinus weirii (‘laminated root rot’ of many fir trees, and a root and butt rot of western red cedar), Heterobasidion annosum (the most damaging root pathogen of coniferous trees in the Northern hemisphere), Thanatephorus cucumeris, which is the teleomorph of Rhizoctonia solani and a very common soil borne pathogen with a great diversity of host plants. Humans don’t escape, either: Filobasidiella neoformans is the teleomorph of Cryptococcus, referred to above as the cause of cryptococcal meningitis in immunocompromised patients.
We turn the tables by eating enormous amounts of hymenomycete fruit bodies: all commercially farmed fungi are hymenomycetes; these include the button mushroom (Agaricus bisporus), oyster mushroom (Pleurotus spp.), shiitake (Lentinula edodes), paddy-straw mushroom (Volvariella volvacea), enokitake (Flammulina velutipes), and shimejitake (Hypsizygus tessulatus). Other edible mushrooms with massive world-wide markets (amounting to billions of dollars annually) are field-collected mycorrhizal species like Cantharellus cibarius (the chanterelle), Boletus edulis (cep, penny bun or porcini), and Tricholoma matsutake (matsutake).
For a long time, the Guinness Book of World Records gave the largest known fruiting body of a fungus as a 160 kg specimen of the polypore Bridgeoporus nobilissimus. This has since been displaced by a very large specimen of Rigidoporus ulmarius (a bracket fungus that grows at the base of the trunks of deciduous trees, usually elm) growing in the Royal Botanic Gardens at Kew which, in the 1999 edition, was recorded as 163 × 140 cm with a circumference of 480 cm. Fruit bodies of this size put the Giant Puffball (Calvatia gigantea) in the shade, as these (edible) hymenomycete fruit bodies average around 10 to 70 cm diameter, although they’ve been known to reach diameters up to 150 cm and weights of 20 kg. The largest, most extensive, and longest-lived mycelium currently known is a mycelium of Armillaria gallica in the Blue Mountains/Malheur National Forest in Eastern Oregon, USA. This one individual fungus covers an area of nearly 900 hectares (3.4 square miles) and is estimated to be more than 2,400 years old; so it constitiutes the largest and oldest living thing known on the planet.
Patterns of evolution of mushroom species across the world have been studied using multigene and genome-based data that contributed to a 5,284-species phylogenetic tree (Varga et al., 2019). To quote these authors:
‘Agaricomycetes started a rapid class-wide radiation in the Jurassic, coinciding with the spread of (sub)tropical coniferous forests and a warming climate. A possible mass extinction, several clade-specific adaptive radiations and morphological diversification of fruiting bodies followed during the Cretaceous and the Paleogene, convergently giving rise to the classic toadstool morphology, with a cap, stalk and gills (pileate-stipitate morphology). This morphology is associated with increased rates of lineage diversification, suggesting it represents a key innovation in the evolution of mushroom-forming fungi. The increase in mushroom diversity started during the Mesozoic-Cenozoic radiation event, an era of humid climate when terrestrial communities dominated by gymnosperms and reptiles were also expanding.’
Lutzoni et al. (2018) had previously found that saprotrophic mushrooms diversified in the Late Paleozoic as forests of seed plants started to dominate the landscape; saying ‘The subsequent diversification and explosive radiation of Agaricomycetes, and eventually of ectomycorrhizal mushrooms, were associated with the evolution of Pinaceae in the Mesozoic, and establishment of angiosperm-dominated biomes in the Cretaceous’.
Updated January, 2020