Resources Box |
|
This outline discussion of pheromones used in mating in fungi is sourced from section 6.1.4 to 6.1.6 in Fungal Morphogenesis (Moore, 1998). Text reference citations have been removed, so refer to the original book for detailed references. The particular point we want you to notice from this discussion is that fungi produce a full chemical spectrum of hormones – terpenoid-, sterol- and peptide-hormones. Just like animals. Chemoattractants in aquatic fungi
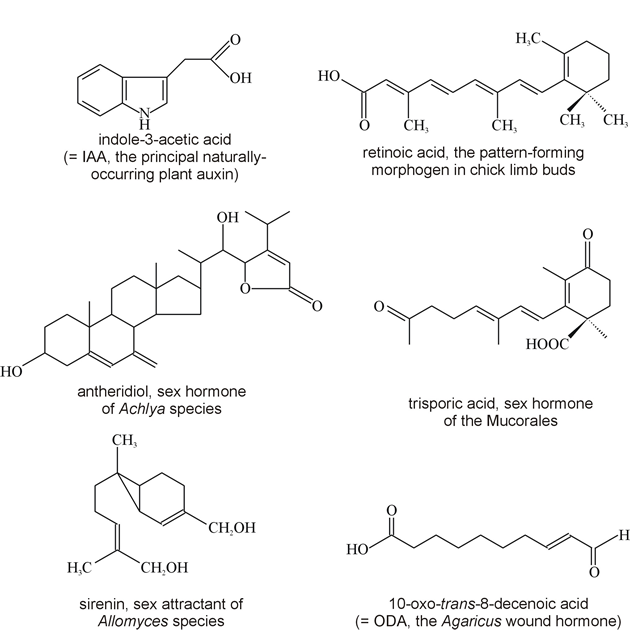
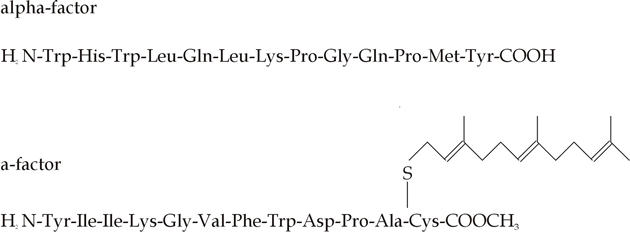
Fig. 1. Chemical structures of some growth factors and hormones. The female sex hormone of another water mould, Achlya (which is not a true fungus but belongs to the Chromista), has also been characterised in some detail. This material, called antheridiol, is a steroid (Fig. 1) the activity of which can be detected by bioassay in 10-11 M solution. The mating sequence reported consisted of: development of antheridial hyphae on the male; production of oogonial initials on the female; growth of antheridial hyphae towards oogonial initials; formation of cross-walls separating oogonia and antheridia; and, after the two made contact, the antheridium grew through a lysed portion of the oogonial wall, after which its own wall was dissolved. Antheridiol is produced continuously by the female and under the influence of the hormone, branches on the putative male thallus which might otherwise grow out as vegetative branches are caused to elongate rapidly and differentiate into antheridia. The male is also induced to excrete a second hormone, oogoniol or hormone B (also a steroid), and it is in response to this that the female initiates oogonial differentiation and amplifies antheridiol levels to those which attract antheridial hyphal growth. There are thus at least two contributors to this hormonal ‘conversation’; the female produces antheridiol but takes up very little itself and does not synthesise oogoniol though it does have a receptor for this hormone. On the other hand, the male makes no antheridiol but is sensitive to it, and one of the responses is to produce oogoniol to which the male is insensitive. Antheridiol and oogoniol are thought to be alternative products of a branched biosynthetic pathway. In the male, antheridiol amplifies that branch of the pathway which leads to oogoniol synthesis and also increases respiration, induces breakdown of glucan reserves in the cytoplasm and triggers de novo synthesis of cellulase. These metabolic changes contribute to processes involved in antheridium initiation, including aggregation of vesicles at the sites where initials develop. It is likely that broadly similar responses are elicited in the female by oogoniol. It is not known how these sterols influence gene regulation but the available data seem to indicate a system very much akin to sterol regulation in animals where the steroid receptors are located in the nucleus and hormone-receptor complexes bind directly to DNA to regulate gene expression. However, antheridiol does not induce changes of this sort and the hormone binding proteins are present in the cytosol, though they are quite similar to other steroid hormone receptors. Chemoattractants in the Mucorales The trisporic acids are synthesised from β-carotene: the molecule is cleaved to retinal, a C-2 fragment is lost, and then there is a series of oxidations. The complete reaction sequence occurs in vitro only when both plus and minus mating types are grown in mixed culture or in an experimental apparatus in which they are separated by a membrane permeable to small molecules. Both mating types have the genetic capacity to produce the enzymes of the complete pathway, but the alleles which determine the mating type repress complementary steps in the later stages of trisporic acid synthesis. Thus, in plus strains synthesis of enzymes needed to form the 4-keto group is repressed by the MT+ allele while enzymes involved in forming the 1-carboxylic acid group are repressed by MT-. Each mating type thus produces a precursor which only the opposite mating type can convert to trisporic acid. The precursors diffuse between the strains and have the status of prohormones which stimulate trisporic acid synthesis. Early steps in the pathway are repressed to a rate-limiting level by a mechanism which allows activation by trisporic acid. When plus and minus strains come together, therefore, the complementary synthesis of trisporic acid consequent on the co-diffusion or the prohormone precursors leads to derepression of the early part of the pathway and amplification of overall trisporic acid synthesis. The increasing gradient of prohormone diffusing from each zygophore induces a chemotropic response. The zygophores can grow towards one another from distances of up to two millimetres. When the zygophores make contact they adhere firmly in a way that implies that mating type-specific and species-specific substances are formed on the zygophore surface. These features are clearly an aspect of the mating type phenotype and are necessary for completion of the mating programme, without adhesion the zygophores continue unproductive extension growth, but the nature of the substances involved is unknown. Chemoattractants in yeasts Mating type factors in the yeast Saccharomyces cerevisiae are responsible for production of peptide hormones (pheromones called α- (= 'alpha') and a-factors; Fig. 2) and pheromone-specific receptors. In animals, pheromones are chemicals emitted into the environment by an organism as a specific signal to another organism, usually of the same species. Effective at minute concentrations, pheromones often have important roles in regulating social behaviour of animals, being used to attract mates, to mark territories, and promote social cohesion in communities. Yeast pheromones were named by analogy to the animal hormones because they organise the mating process. They bind to pheromone receptors on the surface of cells of opposite mating type acting through GTP binding proteins to alter metabolism and to:
Both pheromones cause their target cells to elongate into projections but have no effect on cells of the same mating type or on diploids. In Ustilago maydis, conjugation tube formation in the yeast-like sporidia is induced by mating-type-specific pheromones released by haploid cells. These pheromones are short lipopeptides: 11–15 amino acids with a C-terminal cysteine residue to which a farnesyl group (a 15-C isoprenyl moiety) is attached. The latter makes the pheromone extremely hydrophobic.
Fig. 2. Simplified chemical structures of yeast pheromones. These pheromones prepare the cells for conjugation and contribute to the recognition of different mating types. However, the major step in the recognition of compatible cell types involves macromolecules on the cell surfaces which cause cells to agglutinate. Some of these are constitutive (i.e. cells agglutinate immediately the different clones are mixed) while others are inducible, the cells only acquiring the ability to agglutinate after growth in mixed culture. In both Hansenula wingei and Saccharomyces cerevisiae there is evidence that the molecules directly involved in agglutination – the agglutinins – are glycoproteins. In H. wingei one of the agglutinin components consists of 28 amino acids and about 60 mannose residues. The agglutinins seem to be located on surface filaments external to the cell wall. The function of the agglutinins is to bring cells of opposite mating type together. They do this by virtue of their ability to bind in a complementary manner, the agglutinin of one mating type binding specifically to that produced by the compatible mating type. Following this adhesion of yeast cells by complementary binding, protuberances grow out from the cell walls and when these meet cytoplasmic communication is established by dissolution of the walls. These phenomena are obviously specifically part of the mating process; yet they clearly demonstrate that fungal cells are capable of producing surface glycoproteins which, by a sort of antigen–antibody reaction, can achieve a very specific adhesion. It is exactly this sort of specific cell binding which one might expect to be part of the cell-to-cell communication which contributes to the construction of differentiated multicellular structures. The activity of pheromones is also a model for the way extracellular signals can affect proliferation and differentiation of eukaryotic cells. The signals are detected at the plasma membrane in S. cerevisiae by a family of protein kinases known as the mitogen- or messenger-activated (MAP), or extracellular signal-regulated (ER) protein kinases. This class of enzyme is highly conserved in eukaryotes and as there are at least five physiologically distinct MAP kinase signalling pathways in yeast, it is likely to be involved in transmitting a range of signals through the cytoplasm and into the nucleus to generate changes in metabolism and gene transcription. The pheromone-responsive MAP kinase cascade interacts with a protein required for establishing cell polarity. So spatial information can also be conveyed through regulation of the rearrangement of the actin cytoskeleton. Further reading (much of the detail discussed in these references is dealt with in later Chapters of 21st Century Guidebook to Fungi)Chiu, S.W. & Moore, D. (eds.) (1996). Patterns in Fungal Development, Cambridge University Press: Cambridge, U.K. ISBN: 0521560470. Chiu, S.W. & Moore, D. (1999). Sexual development in higher fungi. Chapter 8 in Molecular Fungal Biology (ed. R. P. Oliver & M. Schweizer), pp. 231-271. Cambridge University Press: Cambridge, U.K. ISBN: 0521561167. Feofilova, E.P. (2006). Heterothallism of mucoraceous fungi: a review of biological implications and uses in biotechnology. Applied Biochemistry and Microbiology, 42: 439-454. DOI: https://doi.org/ 10.1134/S0003683806050012. Moore, D. (1998). Fungal Morphogenesis. Cambridge University Press: New York. 469 pp. SEE chapter 6 Development of Form. ISBN: 0521528577. Moore, D. & Novak Frazer, L. (2002). Essential Fungal Genetics. Springer-Verlag, New York. ISBN: 0387953671. SEE chapter 2 Genome Interactions. CLICK HERE to download the complete text. |
Close the window to return to 21st Century Guidebook to Fungi
This is a Resources Box from the 21st Century Guidebook to Fungi:© David Moore, Geoffrey D. Robson and Anthony P. J. Trinci 2019