
1. Introduction
Prokaryotes have dominated the Earth for the bulk of its history; the last universal common ancestor (LUCA) of all organisms alive today must have emerged close to the start of the Archaean Eon, about 3.8 billion years ago, because some of the oldest microbial fossils are fully differentiated, photosynthetic bacteria (cyanobacteria) found in Western Australian sediments that are 3.5 × 109 years old (Schopf, 1993; Derenne et al., 2008; Boal and Ng, 2010). On the other hand, eukaryotes are generally thought to have appeared no earlier than about 1.5 billion years ago (but see below). So, for at least the first 2 billion years after the origin of life, the living organisms on the planet were prokaryotes together, presumably, with their associated viruses.
The abundant biological activity in the deep ocean volcanic hydrothermal systems of the present day, most of it being dependent on chemosynthesis rather than photosynthesis, has stimulated the widespread appeal of theories of a ‘deep-hot’ origin of life (Wächtershäuser, 2006; Alpermann et al., 2010). This implies that the pioneer organisms were hyperthermophiles (Stetter, 2006), a notion which builds upon Carl Woese’s conclusion that the three domains, now called Eubacteria, Archaea and Eukaryota diverged from LUCA (Woese, 1987; Woese et al., 1990). Emerging from these arguments we have what might be called a conventional, or ‘textbook’ phylogenetic tree of life (for example see Moore et al., 2011; p. 24). Unfortunately, gene trees are ambiguous and the root of the universal tree of life remains controversial (Penny and Poole, 1999).
The most complete reworking of the tree of life is that recently published by Tom Cavalier-Smith (Cavalier-Smith, 2006, 2010a & b). Cavalier-Smith’s approach is to integrate palaeontology with comparative study of present day organisms, emphasising key steps in molecular and cellular evolution. Cavalier-Smith (2010a) identifies five successive kinds of cell: (i) The first cells were negibacteria, with cells bounded by two acyl ester phospholipid membranes, divided into the primitive anaerobic Eobacteria without lipopolysaccharide in the outer membrane and more advanced Glycobacteria with lipolysaccharide (e.g. oxygenic Cyanobacteria and Proteobacteria); (ii) unibacteria, with one bounding and no internal membranes, divided into desiccation-resistant posibacteria, ancestors of eukaryotes, and archaebacteria as the youngest bacterial phylum and a sister group (not an ancestor) of eukaryotes; (iii) eukaryotes with endomembranes and mitochondria, (eukaryotes plus archaebacteria make up the neomura); (iv) plants with chloroplasts; (v) chromists with plastids inside the rough endoplasmic reticulum.
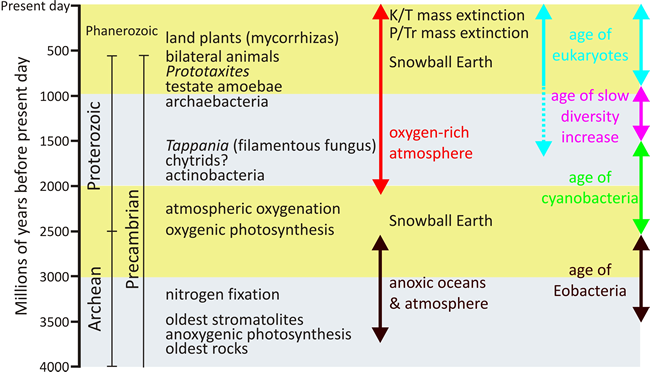 |
| Fig. 1. A geological timescale covering from the time of the oldest rocks (3.8 billion years ago) to the present day, highlighting major geological and evolutionary events and features mentioned in the text, including my interpretation of Cavalier-Smith’s four ages of life at extreme right. Note that Cavalier-Smith’s age of eukaryotes starts 850-800 million years ago, but as I interpret Tappania fossils to be fully differentiated sclerotia of filamentous fungi I place the origin of stem eukaryotes somewhere between 2000 and 1500 million years ago. Modified and redrawn from Cavalier-Smith, 2010a. |
These types of cell are placed into four ages of life as follows (Cavalier-Smith, 2010a; see Figs 1 and 3): (i) the age of Eobacteria, an anaerobic phase in which photosynthetic non-sulphur bacteria (and before them extinct stem negibacteria) were the major primary producers. Exclusively anaerobic life probably persisted from about 3.5 billion years ago to just under 2.5 billion years ago (the best date for the origin of photosystem II and start of oxygenic photosynthesis). (ii) The age of cyanobacteria (about 2.5-1.5 billion years ago) during which cyanobacteria were the major primary producers (and are now the dominant morphological fossils). (iii) An age of slow diversity increase (1.5-0.85 billion years ago) features increasing morphological complexity and colonisation of continental surfaces by both Cyanobacteria and, following loss of the outer membrane, Posibacteria and the actinomycete Actinobacteria; the latter displaying the greatest morphological complexity. Some of the largest microfossils from this part of the middle Proterozoic have been attributed to eukaryotic algae, filamentous fungi or stem eukaryotes of undefined affinity (Javaux et al., 2001), though Cavalier-Smith is sceptical of all such fossil identifications in this period. (iv) The age of eukaryotes and obvious macroorganisms (850-800 million years ago to the present).
Cavalier-Smith (2006) argues that eukaryotes derived from an actinobacterial ancestor on the grounds (among others) that current Actinobacteria are the only eubacteria having phosphatidylinositol, which is one of the most important eukaryote phospholipids, required for eukaryote specific cell signalling. “...Thus, eukaryote membrane lipids probably came vertically from an actinobacterial ancestor, archaebacterial lipids originating in their [last common ancestor] after it diverged from eukaryotes.” A further aspect of this argument is that shortly after eukaryotes and archaebacteria diverged, the latter group colonised hot, acid environments by evolving the ancestrally hyperthermophilic archaebacteria and later, one archaebacterial lineage evolved biological methanogenesis; (Cavalier-Smith, 2006 pp. 977-978).
Updated December 7, 2016