解讀真菌的形態發育
趙紹惠 和 大衛摩亞
Department of Biology, The Chinese University of Hong Kong, Shatin, N. T., Hong Kong SAR, China
中國香港新界沙田 香港中文大學生物系
&
School of Biological Sciences, University of Manchester, Manchester M13 9PT, U. K.
英國曼徹斯特市 曼徹斯特大學生物科學學院
第六章:細胞分化和形態發育的遺傳機制
Genetic Basis for Cell Differentiation & Fungal
Morphogenesis
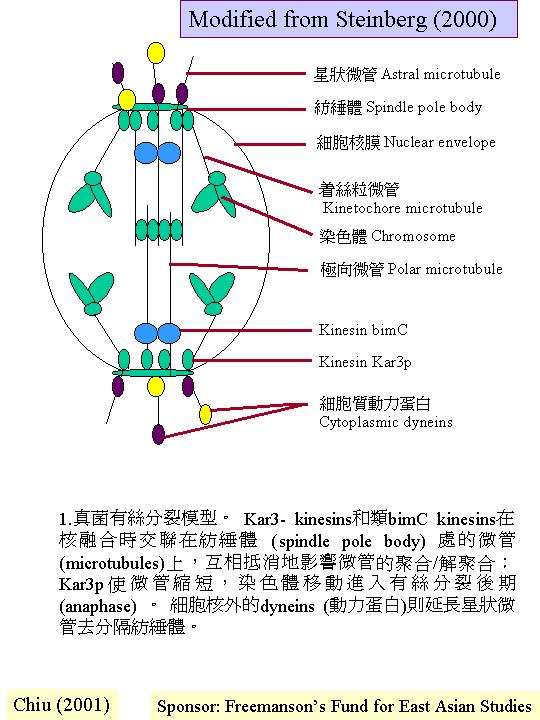
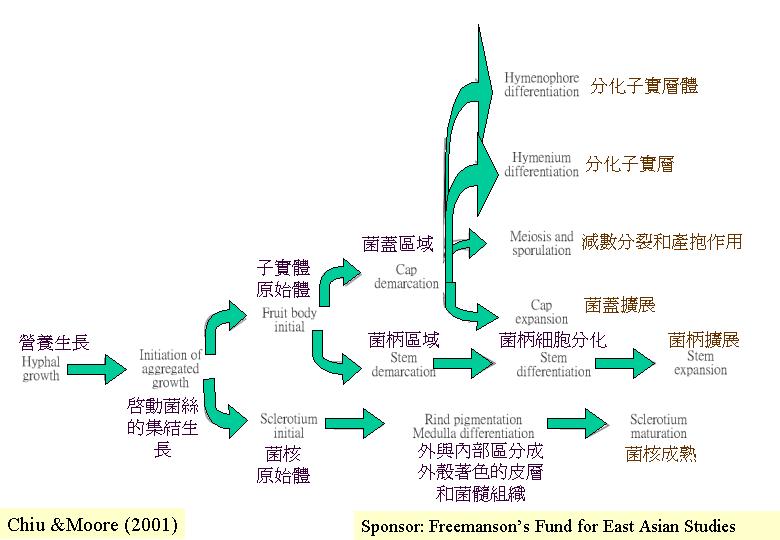
真菌相對其他真核生物有較小的基因組(Clutterbuck, 1995b)。真菌細胞核和染色體都比較小、多變化 (meiosis),因此,在普通光學顯微鏡下並不清晰,阻礙了真菌細胞學的發展。真菌有絲分裂是在細胞核內進行的 (intranuclear mitosis) (mitosis ) ;細胞核膜在細胞週期均保持完整。此外,真菌的主要生活期常是單倍體的 (haploid) ,或獨特的雙核期 (N + N) ,雙倍體 (diploidy) 就在減數分裂 (meiosis)前短暫出現。雙核的菌絲體可作營養體繁殖下去,直至適合的內和外條件下,就製造有性或無性的多細胞結構。從分裂面的位置,可見真菌的特徵:菌絲的細胞分裂而形成橫壁 (橫隔, septum) 與管細胞軸成直角;真正的縱壁構造只是在銀耳目等的擔子上找到。
真菌的細胞分化與細胞核的數目和倍體沒有特定的關係。雙孢蘑菇(Agaricus bisporus) 的營養菌絲體有六—廿個細胞核 (Colson,
1953; Kligman, 1943) ,擔子果細胞平均有六個核 (Evans, 1959) ,菌柄細胞的可達32個 (Murakami & Takemaru, 1980) 。灰蓋鬼傘 (Coprinus cinereus) 是一雙核體,但菌柄細胞是多核的,數目達 156之多 (Stephenson
& Gooday, 1984; Gooday, 1985)。最特別是蜜環菌 (Armillaria)
,其營養菌絲體是雙倍體的,但其生活史上有可能出現二次或以上的單倍化的情況 (haploidization) 。同宗結合 (homothallic) 的菌株和異宗結合 (heterothallic) 的菌株就出現不同的核循環
(Chiu et al., 1993) 。
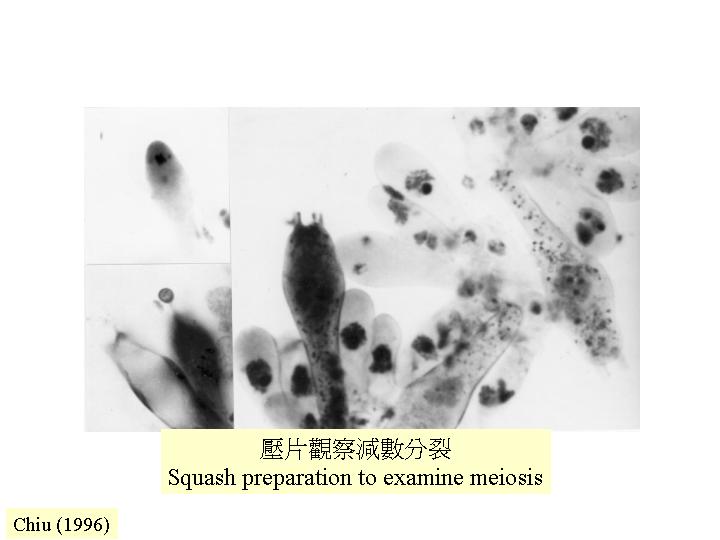
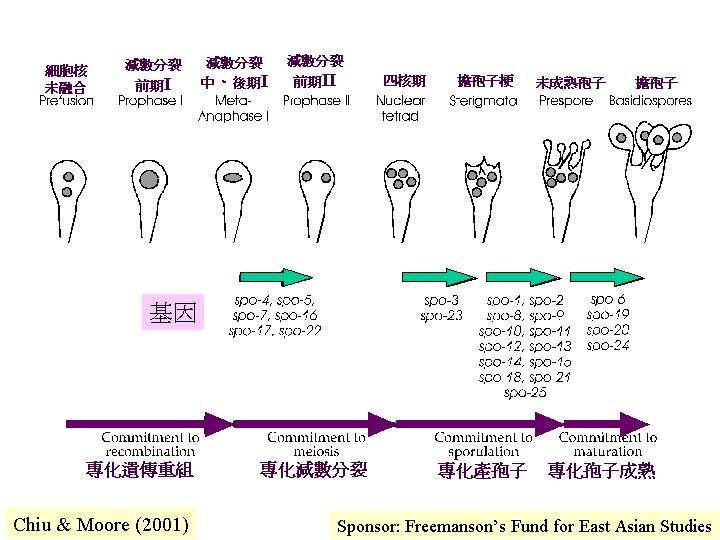
減數分裂細胞 (meiocyte)
真核的 DNA複制需要 α DNA 聚合 (polymerase) 作用:領導聚核酸鏈 (leading strand)的開始合成和落後聚核酸鏈 (lagging strand) 的合成。在雞胚胎研究發現:當靜止細胞被激活時,α DNA 聚合的mRNA水平增加,轉譯速率和活性都增加。進入細胞分化時,α DNA 聚合快速消失。因此,α DNA 聚合不是細胞週期調控的。灰蓋鬼傘的減數分裂前的DNA複制 (pre-meiotic replication) 就必需α DNA 聚合,出現在減數分裂前期 (premeiotic) 至減數分裂早期 (early meiotic prophase) ,進入減數分裂後,α DNA 聚合-引發複合物 (alpha DNA polymerase–primase complex) 分離,只殘留催化子單元 (catalytic subunit) 。
而減數分裂包括以下步驟:聯會
(synapsis) 、減數分裂的基因重組 (meiotic recombination) 和分離 (segregation) (Hawley &
Arbel, 1993; Moens, 1994)。減數分裂前的 DNA複制和形成聯合複合體 (synaptonemal complex) 都是獨立事件
(Kanda et al., 1989a, 1990; Pukkila, 1994)。而染色體配對和聯會的機制和時間都有分別 (Padmore et
al., 1991) 。進行聯會 (synapsis) 並不要求 DNA同源性 (DNA homology) (Hawley & Arbel,
1993; Pukkila, 1994) 。在前期一 (prophase I),染色體配對,然後聯合複合體出現,隨後DNA鏈斷裂讓減數分裂的基因重組可發生。減數分裂的基因重組
(meiotic recombination) 中,異源雙鏈DNA (heteroduplex DNA, hDNA) 會出現,這雙鏈DNA可有一或多個錯配的鹼基配對。DNA-
DNA的重組不足以引起染色體分離 (disjunction) (Hawley & Arbel, 1993; Pukkila, 1994) ,但染色叉
(chiasmata) 就專責平衡染色體上絲粒所受的拉力,去確保同源染色體在減數分裂中期 I 時的位置。減數分裂的基因重組率與有絲分裂的,高出100 – 1,000 倍(Hawley & Arbel, 1993;Moens, 1994)。減數分裂 I 為核的倍數體減半,減數分裂 II為平衡分裂,真核的減數分裂與其他真核生物無異,只是,細胞核膜均保持完整
(intranuclear divisions)。
第五章(Chapter 5)論述擔子的分化途徑,其中灰蓋鬼傘的
rad (radiation-sensitive mutants) 基因突變種就不但無法完成減數分裂而且影響產孢程序 (Chiu, 1988;
Bourne et al., 1996) 。最近,在蠔菇 (Pleurotus ostreatus) 就找到一DNA序列
(DMC1 homolog) 屬 RecA-like (recombination, 基因重組; RAD51 組;) 的基因
(Mikosch et al., 2001)。細菌大腸幹菌的RecA負責DNA鏈的配對和交換 (pairing and strand exchange) ,而釀酒酵母 (Saccharomyces
cerevisiae) 的 RAD51基因與DNA雙鏈斷開的修復和重組有關,鬼傘的
CoLIM15和 RAD51 基因就在減數分裂或DNA受損時誘導的 (inducible) 。酵母
DMC1或RAD51基因若有突變 (鬼傘的RAD基因突變種都是) ,則不完成減數分裂前期,和不產孢子,但RAD51基因突變菌株對DNA破壞劑 (化學品和輻射) 敏感,DMC1基因突變菌株卻沒有這敏感反應,DMC1基因轉錄產物只在蠔菇菌褶找到,營養菌絲和擔子果的其他構造都找不到 (Mikosch
et al., 2001)。
文獻上報導:有少數人對食用菌孢子過敏
(allergic) ,因此,有些育種者 (breeders) 都希望找到不產孢子 (sporeless) 的菌株,而有少數菇農在長期接觸下,而變成對大型食用菌孢子過敏,(如:估計蠔菇一天產
6 x 10 8 孢子/ g 子實體,工業生產時空氣中的孢子量會相當高),因此,菇場加裝孢子收集器去處理排出的廢氣,和菇農工作時帶面罩,都則保障員工健康的措施。不過,靈芝孢子的售價遠高金價
(HK$16,000/kg(= US$2,000/kg) in 1999),如何提高產量和減少生產所需時間便變得重要。
兩株釀酒酵母 (Saccharomyces cerevisiae) 在(pulsed field gel electrophoresis)脈場凝膠電泳下顯示染色體長度多態性 (chromosomal length polymorphism) electrokaryogram; Chiu & Moore, 1999)。脈場凝膠電泳技術 (pulsed field gel electrophoresis) 和DNA 雜交 (hybridisation) 技術的發展令我們發現:真菌發育分化時有基因增殖 (gene amplification) (Pukkila & Skrzynia, 1993) 、不同菌株表現染色體多態性和有些染色體的缺失並不影響成活率 (Zolan et al., 1994; Carvalho et al., 1995) 。由此可見真菌基因組的彈性 (genome plasticity) (Tooley & Carras, 1992; Martin, 1995) 。雖是這樣,人工創造的三倍體和四倍體灰蓋鬼傘 (Coprinus cinereus) 的擔子果都出現畸型發育 (Murakami & Takemaru, 1984) 。而釀酒酵母 (Saccharomyces cerevisiae) 的三倍體和四倍體就出現三鏈聯會 (triple synapsis) 和聯會的雙鏈 (synapsed bivalents) (Loidl, 1995) 。
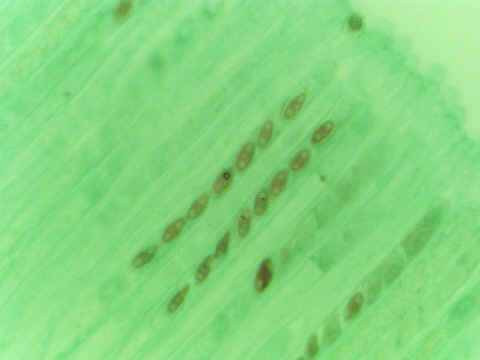
Chiu
(1996) 指出擔子菌的減數分裂後的有絲分裂
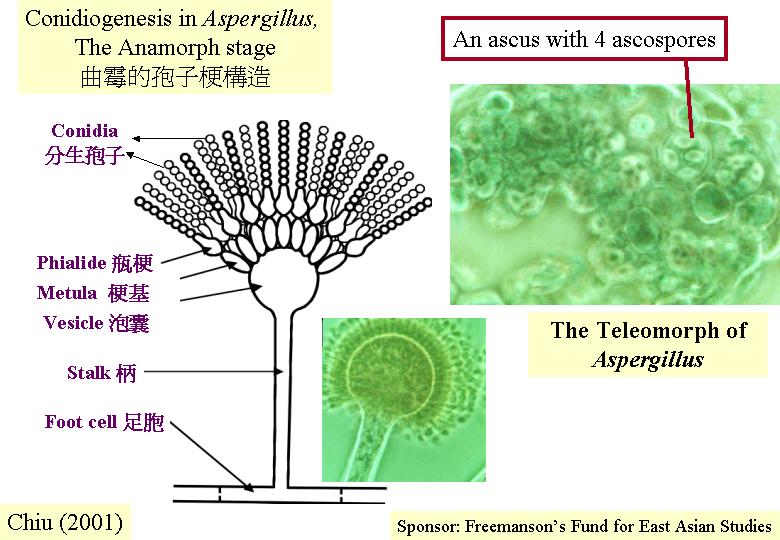
(postmeiotic meiosis) 有多種形式:沒有有絲分裂、有絲分裂在擔子或擔子梗或擔孢子內進行。許多子囊菌都進行一次的減數分裂後的有絲分裂,結果,子囊有八個子囊孢子 (ascus with 8
ascospores)。糞盤菌(Ascobolus
immerses) 、 粗糙孢菌 (Neurospora crassa) 和糞生糞殼 (Sordaria
fimicola) 製造有規律的四分體 (ordered tetrads) ,因為減數分裂的子核位置反映第一或第二減數分裂的分離
(segregation)。
親和系統與交配型基因
Incompatibility &
Mating type Genes
一個單核孢子萌發
(nuclear migrtion
to germtubes )形成一同核菌絲體 (homokaryon),透過營養親和系統
(vegetative compatibility, 體質不親和系統, somatic incompatibility) 和性不親和系統 (sexual
incompatibility) 的控制,兩個相遇的同核菌絲體可融合成一異核菌絲體 (heterokaryon) (to develop) ,或拱固各自的地盤,形成各自的領域 (territory) ,又或出現競爭現象,侵食其中一方。動物藉外激素來吸引週遭環境中的同種生物, 在極微濃度下, 外激素有效地管理動物的社群行為: 吸引配偶、標記領土範圍, 而且促進結合形成一社區。在真菌,就算是兩個相遇的異核菌絲體,也可發生地盤性的競爭,或重新交換細胞核 (nuclear exchange),當中或許發生營養性的基因重組 (somatic recombination),甚至形成多個異核菌絲的共同體
(clone)。蜜環菌 (Armillaria gallica) 的地盤可有43公頃的面積,約有1, 500歲,相信是地球最老和最大的生物
(Smith et al., 1992) 。
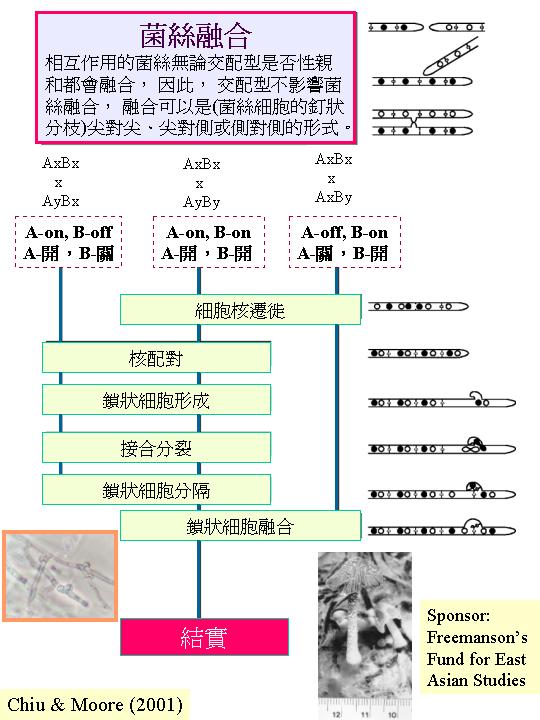
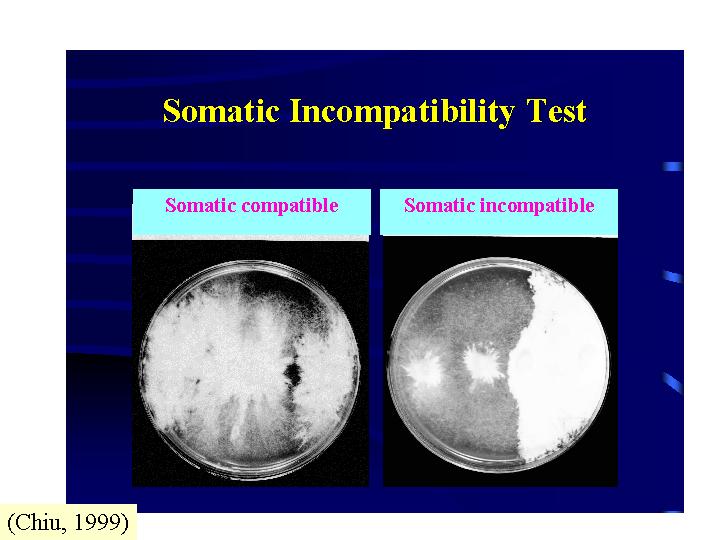
真菌的性生活源於兩性親和的同核菌絲體,相遇、菌絲融合、細胞質融合、細胞核交換 (mating
type control)。如果兩相遇的群落擁有相同的基因型,營養親和系統 (vegetative compatibility, 體質不親和系統,
somatic incompatibility, 異核體不親和系統, heterokaryon
incompatibility) 則專責防止菌絲融合、細胞質融合 (individualism)。營養親和系統是一多基因的系統。但若個體可交換細胞核,性親和系統
(sexual compatibility, 交配系統mating system) 控制性生活 (Chiu & Moore, 1999)。
不是所有真菌都進行交配
(mating),才可有減數分裂和產孢程序,例子如: Coprinus bilantus。但在 Neurospora和 Sordaria 的一些同宗結合的 (homothallic) 種株中,有找到交配型因子
(mating factor) 的 DNA序列 ,所以,這可能代表交配型因子的座位上
(chromosomal locus)有操控性繁殖的或結實的結構基因。又或者生物向有性繁殖演進,去提高產生遺傳變異的效能,但環境因素令某些生物返回自我受精,固保留交配型因子的
DNA序列。交配型因子 (mating factor) 是令個體與「非我」結合。真菌屬微生物,進行無性繁殖去增加個體數量,所以,在週遭遇起「自我」的機會是有的。真菌交配型因子的遺傳糸統可有:
i.
一個交配型因子 (unifactorial) 兩個等位基因yeast sex control、
ii. 一個交配型因子多個等位基因、
iii. 甚至兩個交配型因子
(bifactorial) ,而每一座位都出現多個等位基因 (2
genetic loci, each has multiple alleles) fungal sex control。
這多等位基因現象
(multiple allelism) 是將交配成功率由 50%提高。另外,同一交配型因子上有兩個以上同功能的連鎖基因
(lined genes),可能代表「基因重複製造」 (gene
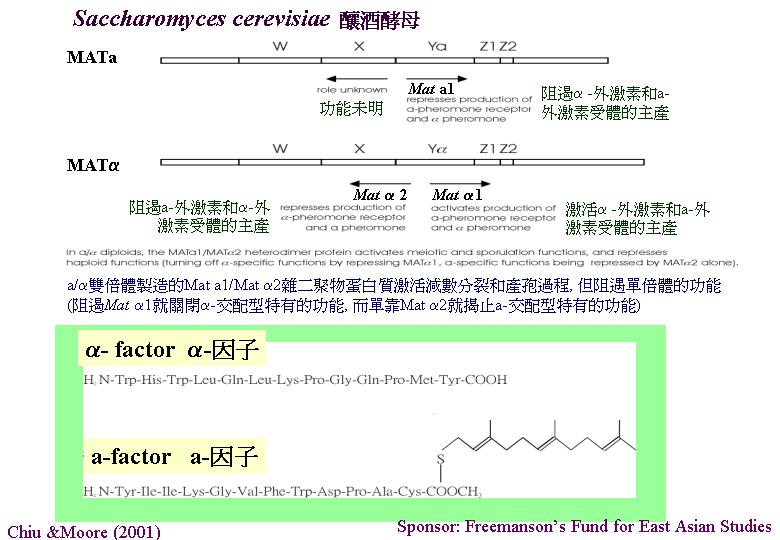
duplication) 的現象 ,亦提高交配成功率。 釀酒酵母 (Saccharomyces cerevisiae)
的三個遺傳座位HML、 MAT 和 HMR交配型座位都在同一染色體上,釀酒酵母進行變換交配型
(mating type switching) 模式。
玉米黑粉菌 (Ustilago maydis) 的單倍體細胞以酵母形式出現,而且是腐生的 (saprotrophic) ,不能感染植物的 。只有經交配而形成的雙核體 (dikaryon) ,轉形成菌絲體,而且必須感染植物玉蜀黍 (Zea
mays) 才能維持生長和進行有性繁殖,製造 teliospores (heterobasidia) (Kämper et al., 1994)。受感染的玉蜀黍出現的病徵包括:莖、葉、花序腫瘤,因為玉米黑粉菌分泌植物激素
auxin (indole-3-acetic acid; IAA) (Bőlker, 2001)。
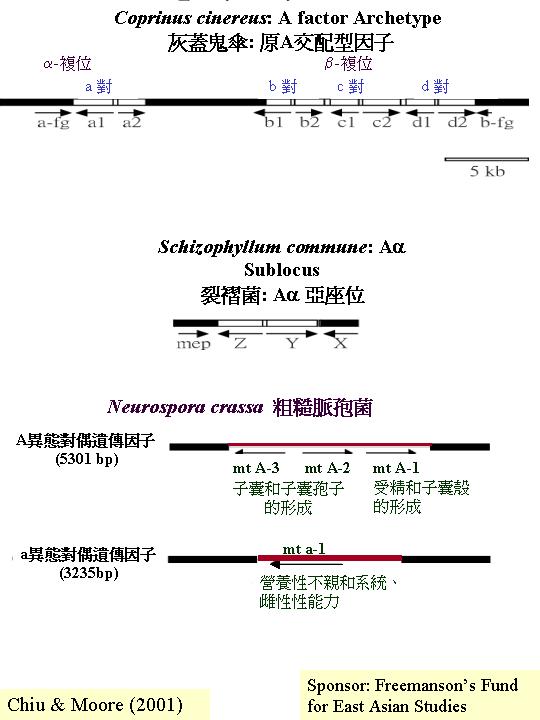
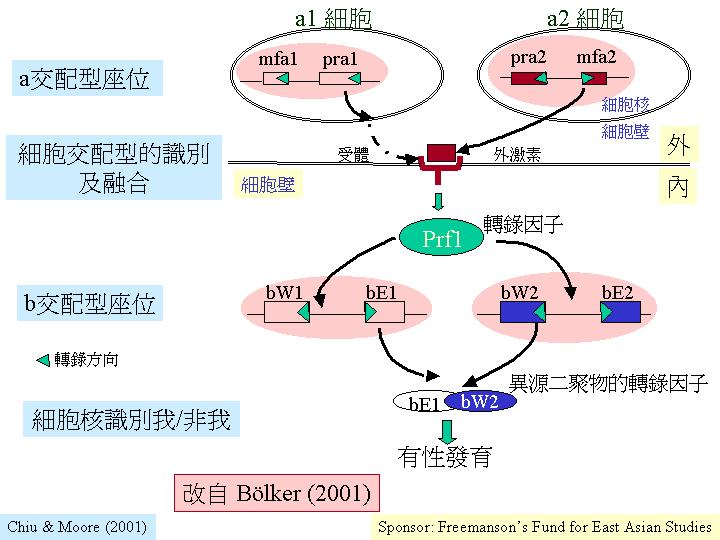
玉米黑粉菌 (Ustilago maydis) 的交配型因子a 和 b的構造 (Ustilago
sex control)。 a座位上的同位基因擁在一交配型特有的可變DNA序列位置
(a1為4500 基對,a2為8000基對), 而保守的DNA序列 (塗黑處)是兩個交配基因 mfa 和 pra, b座位有兩個閱讀框 (reading frame) bW 和 bE,而轉錄成的多,在保守區域
(domain) (塗黑處) 有90%以上的相同序列,而在可變區域(domain)則降至60-90%。可見到轉錄因子是重要的調控系統
(Bőlker, 2001) 。
總論:交配型基因作為「總管」,製造轉錄因子 (transcriptional factors) 去調控 (激活和阻遏) 其他基因,影響性發育,包括:外觀、獨特的細胞-細胞相互作用、細胞形態和行為,開啟新的發育途徑。
交配型基因的主要功能特徵,包括:
i.
有多個閱讀框。
ii.
遺傳座位是固定的,但對位基因DNA序列差異大,轉錄不同產物,固稱idiomorphs。
iii.
部分真菌表現交配型變換。
iv.
部分轉錄物是轉錄因子,組合的異源異聚物,調控其他基因。
v.
部分轉錄物是外激素、外激素戴體,調控細胞與細胞的相互作用。
可見交配型基因並不直接參予真菌的形態發育。
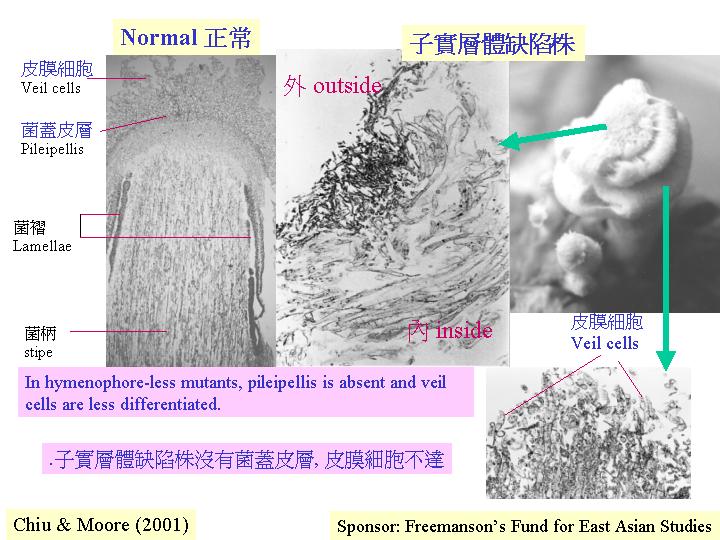
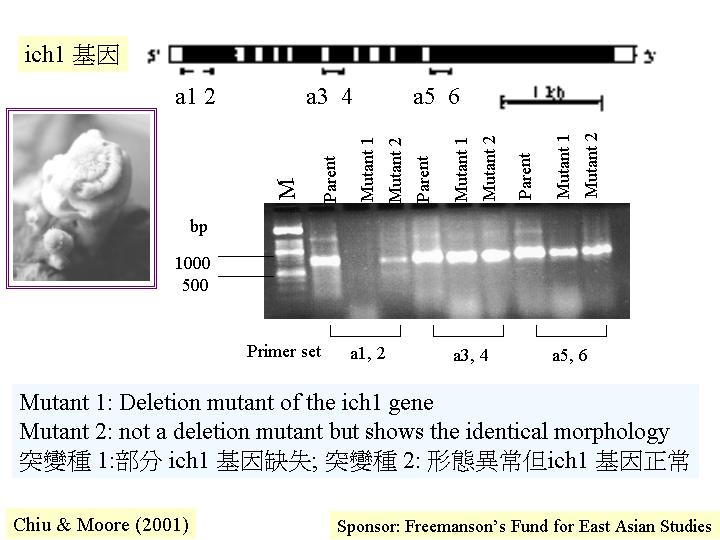
最近在灰蓋鬼傘 (Coprinus
cinereus) 獨立地找到兩株轉錄因子的基因缺陷菌株 (deletion
mutants) (Kamada et al., 1998; Chiu & Moore, 2000) ,同一表現徵兆就是缺乏子實層體
(hymenophore-less) 的畸型擔子果而非菌蓋 (pileus)
(hymenophoreless
mutant ) (detection of
gene defect) (Chiu & Moore, 2000) 。
菌絲的無性繁殖:製造分生孢子
其實,我們對真菌形態發育的認識非常有限,不過,這裡作一個小的總結:真菌形態發育包括反應能力 (勝任性competence) 、誘發 (induction) 和改變
(change) 。如:構巢曲霉 (Aspergillus nidulans)
的無性繁殖有三個步驟:細胞生長到達勝任作分化期,誘發發育途徑和進行製造分生孢子,菌絲形態改變成分生孢子梗 Aspergillus
1。
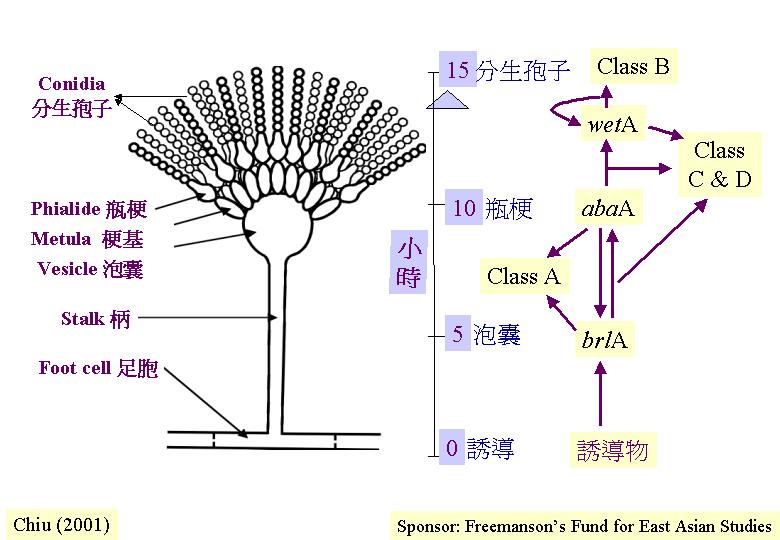
分生孢子梗
(conidiophore) 與營養菌絲的分別有三:分生孢子梗由厚壁的基部足胞 (foot cell) 延伸而來,足胞有兩層細胞壁,外壁與菌絲體其作細胞的相連,內壁連與分生孢子梗的。分生孢子梗的直徑比菌絲體其作細胞的長。還有,分生孢子梗少分枝,表現有限生長
(determinate growth) 。15 - 30分鐘的紅光照射激活分生孢子生產途徑 (Asp2)。
經傳統的遺傳手段 (誘變、選擇和分柝突變種) 可建立基本的遺傳調控構巢曲霉分生孢子梗發育的途徑
(Clutterbuck, 1977, 1978; Champe et al., 1981; Timberlake et al.,
1983; Gwynne & Timberlake, 1984; Timberlake & Hamer, 1986; Timberlake,
1987) 。約有 300 基因座 (loci) 參予分生孢子梗的發育,1,200
新的信使RNA (mRNAs) 出現在分生孢子梗的組織上,另
200 mRNAs 在分生孢子上 (Martinelli & Clutterbuck,
1971; Timberlake, 1980, 1986) 。
大部分的不產分生孢子誘變種 (aconidial mutant) (83%)在分生孢子梗出現前期已有缺陷,15%影響分生孢子的萌發或產生色素 (其中的 85% 在營養生長也有缺陷),只有約 2%直接與分生孢子梗的生長和發育有關 (Martinelli & Clutterbuck, 1971) 。其中,剛毛基因 (bristle gene, brlA) 控制瓶梗 (phialide)
、梗基 (metula)
和泡囊 (v
esicle) 的形成,而珠細胞基因 (abacus gen, abaA) 透過控制瓶梗出芽階段將分生孢子改為珠狀細胞的模式 (Clutterbuck, 1969; Martinelli, 1979; Johnstone et al., 1985; Timberlake et al., 1985; Boylan et al., 1987)。這兩個基因座各自有30 多個突變對偶基因 (mutant alleles) 。而濕基因 (wetA) 在分生孢子成熟初期作用,使分生孢子不產色素和乏疏水性,更在數小時內自溶 (Clutterbuck, 1969; Gwynne et al., 1984) 。這三個基因的表達時序為:brlA→ abaA → wetA (Martinelli, 1979; Zimmerman et al., 1980; Boylan et al., 1987)。其分子調控途徑以圖 5.12 和 5.13示 (Timberlake, 1990, 1993; Han et al., 1993; Timberlake & Clutterbuck, 1993; Lee & Adams, 1994) 。其中,brlA產物為鋅指多 (zinc finger polypeptide) ,許有轉錄因子都有鋅指的次等構造,相信負責特定DNA序列的結合,為一正轉錄調節器 (Aspergillus 1)。
許多brlA基因突變菌株只表現部分表徵的失活,因而估計brlA基因產物對不同的靶基因
(target genes) 有不同的結合能力。實則 brlA 座位有兩個重的轉錄單位:brlAα 和brlAβ。構巢曲霉brlA 座位的結構。brlAα的
mRNA由第八染色體上的BrlA 基因轉錄出來,而 brlAα序列是單一的基因外區
(exon) ,編碼 (encoding)成一 Cys2‑His2
的鋅指多 , 但 brlAβ 序列有一基因內區(intron) , brlAβ編碼的多另有一由 23個氨基末端 (amino‑terminal
residues) , 和 轉錄本上游有一短的開放閱讀框 (open reading
frame, ORF) [初轉錄本有一ATG起始的閱讀框為41個氨基末端 (稱μORF)], 相信是調節brlAβ轉譯的 (Timberlake, 1993)。
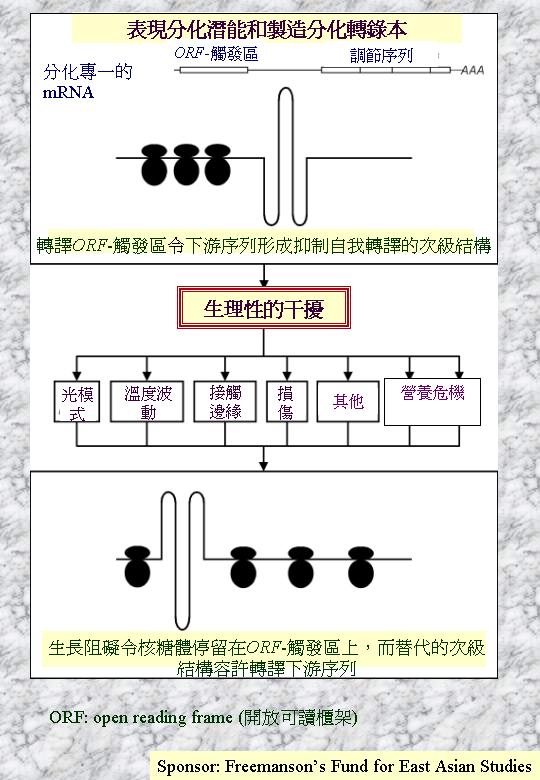
Han
et al. (1993) 透過分子生物學的分析,發現brlAβ轉錄本在營養菌絲可測到,但受到開放閱讀框 (μORF) 起始的轉譯抑制下游
(BrlA) 閱讀框的轉譯;因此,分化定型 (competent) 的菌絲本可發育分生孢子梗,但遇到轉譯抑制 (translational repression) ,保持營養生長。Timberlake (1993) 則指出轉譯觸發 (translational
trigger) 這個理論 (translational
triggering) :若brlAβ轉錄本可克服
μORF的轉譯抑制,其多 (BrlA) 則激活形成分生孢子,其中營養狀 (如缺氮的環境因素) 可降低氨-轉RNA
(aminoacyl-tRNA) 量,影響 μORF的轉譯。
BrlA 的激活只是發育分生孢子梗的第一步,其產物激活專一形成分生孢子的基因如:rodA
(編碼為分生孢子細胞壁的疏水蛋白 (hydrophibin)) 、 yA [編碼為 p-二酚氧化 (蟲漆) 使黃孢子轉綠] ,和 abaA (另一調控基因生產轉錄因子) (直接激活或聯同
medA激活) 。brlA和 abaA基因是相互的激活因子。而abaA產物也激活在瓶梗的wetA
這調控基因,wetA是自我調控表達的,驅使分生孢子在物理上或細胞學上與瓶梗分開。
Mirabito
et al. (1989) 根據調控基因突變菌株的轉錄本質量情況,將不受調控的發育專一基因分成四類:第一類基因由 brlA
和/或 abaA激活,但不受wetA 影響,作用時間為早期發育。第二類基因由wetA激活,但不受brlA 或 abaA影響,作用時間為晚期 (分生孢子的發育) 。第三和第四類基因要求三個調控基因的共同作用下,才表達瓶梗特有的功能 。
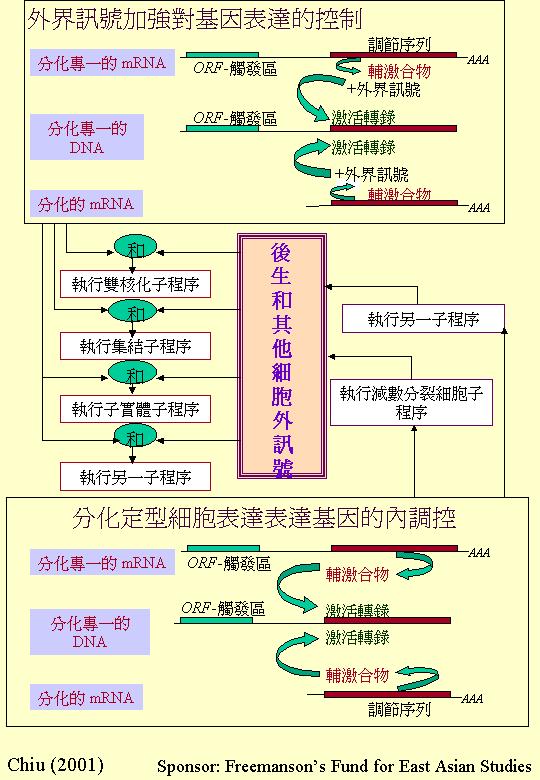
構巢曲霉分生孢子梗發育的遺傳調控顯示:真菌組織進入分化途徑靠轉譯觸發
(translational triggering) 作決定機制的 (translational
triggering)。這轉譯觸發一方面將達致定型分化 (分化反應能力,
development of competence) 與發育連接,另一方面與環境誘導 (initiation) 發育聯繫起來。再者,發育途徑的重點調控機制包括:相互激活
(reciprocal activation) 、反饋激活 (feedback activation) 和自我調控
(autoregulation) 使啟動後的發育程序不再受外界環境因素影響,這便是 Timberlake
(1993) 提出的反饋固定 (feedbackfixation) ,形成經典胚胎學的發育決定
(developmental determination) 的觀念。此外,構巢曲霉分生孢子梗發育中有許多結構性基因 (structural
genes),在其他發育途徑都有表達,正說明發育專一性由調控因子 (regulators) 控制,這樣,真核生物的發育就合乎經濟效益的原則。
真菌形態發育是連串基因操控的子程序而連接的過程。而後遺傳(epigenetics) 只影響基因表達。真菌菌絲有許多形態發育受制於緘默基因 (silencing genes) (Irelan
& Selker, 1996) 。一類的可逆性 (reversible) 轉錄緘默機制 (transcriptional silencing) 就是將重複 DNA 序列 (repetitive DNA sequences)上的胞嘧啶 (cytosine) 甲基化 (methylation) ,如發生在 Ascobolus immerses 和灰蓋鬼傘 (Coprinus cinereus) 減數分裂前 (Meyer, 1996) ,及二型酵母 (dimorphic yeast) 在轉形態時都有發生 (Reyna Lopez et al.,
1997) 。但是,這機制並不普遍和沒足夠証據支持,固不能成為形態發育的網絡因子。
多細胞的性器官
真菌的形態發育的基因是受制於轉錄因子的
(de Vries et al., 1980; Huang & Staples, 1982; Lee & Dean, 1993;
Schuren et al., 1993) 。而轉譯觸發是構巢曲霉分生孢子梗發育的遺傳調控,使外源因子傳訊到形態發育去。
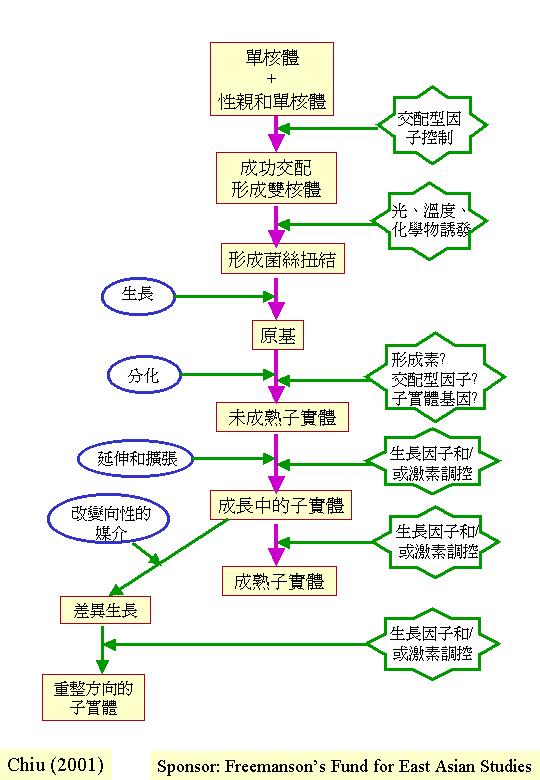
在擔子菌中,單核體產子實體的情況不一定罕見;
27% Sistotrema 的單核體菌株 (Ullrich, 1973) 、7%裂褶菌 (Schizophyllum
commune) 單核體菌株 (Raper & Krongelb, 1958) 和 16株中的一株灰蓋鬼傘(Coprinus cinereus) 單核體
(Uno & Ishikawa, 1971) 才表現結實能力。而且,許多這類的擔子果都是形態不正常、有缺陷、不育等,有關基因部分會在雙核體擔子果表達
(Stahl & Esser, 1976; Esser & Meinhardt, 1977; Esser et al.,
1979) 。Horton & Raper (1991) 利用轉化實驗
(transformation) 發現導致裂褶菌 (Schizophyllum commune) 單核體結實的
DNA段,但有關機制仍未清晰。最有趣是:Verrinder Gibbins & Lu (1984) 發現灰蓋鬼傘(Coprinus
cinereus) 單核體在營養缺乏下出現雙核體結實,估計是營養缺乏誘發其交配型變換,可惜作者沒跟進作這類擔子果後代遺傳的分析。
Takemaru
& Kamada (1971, 1972) 對灰蓋鬼傘 (Coprinus cinereus,舊稱謂是 C. macrorhizus) 的雙核體結實作詳細遺傳分析,利用誘變劑處理雙核體菌絲,從10,
641株生存的菌絲體中,獲得1,591株擔子果發育突變種,根據擔子果表現型可分為:
1. 無扭結
(knotless) :沒有形成菌絲扭結。
2. 無原基
(primordiumless) :沒有形成擔子果原基。
3. 不成熟
(maturationless) :擔子果原基形成後但不成熟。
4. 不延伸
(elongationless) :擔子果菌蓋發育正常但菌柄不延伸。
5. 不開展
(expansionless) :擔子果菌柄延伸但菌蓋不開展。
6. 缺孢子
(sporeless) :擔子果看來正常但擔孢子無或缺。
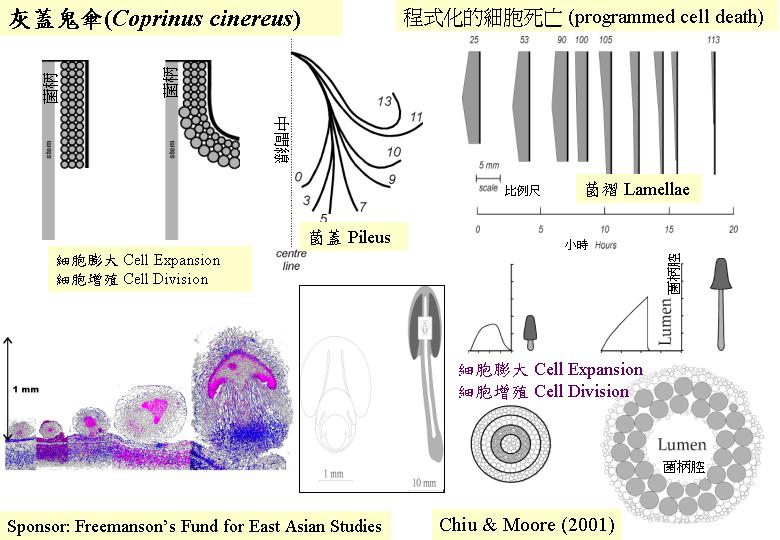
這些雙核體擔子果發育突變種全是顯性遺傳的,這些突變種讓我們研究和了解菌柄延伸
(Kamada & Takemaru, 1977a, b, 1983) 、擔孢子形成過程等 (Miyake et
al., 1980a, b)。
當交配型因子的雙突變種 (AmutBmut
strains) 人工育種出現後 (Swamy et al., 1984) ,對擔子果生長發育的研究提供方便,因為這些單倍體的表現型為同核的雙核體,而且結實正常,因為是同核體,故隱性基因亦可表達出來,而廣被應用。
對灰蓋鬼傘擔子果不延伸
(elongationless) 的發育誘變株作的研究 (Muraguchi & Kamada, 2000)發現:微粒體的細胞色素
P450系統 (microsomal cytochrome P450 system (CYP502) ) 表現組成型活性
(constitutively expressed) ,但顯性(dominant) 的缺失突株
(deletion mutant;有關基因名eln2) 就引起擔子果不延伸的表現型。
這發育缺陷是由於菌柄細胞數量不足,而不是細胞大小的問題。從
DNA序列而推斷的氨基酸序列可推斷為新一族的cytochrome P450 family
(http://www.icgeb.trieste.it/p450)。而建築的雙孢蘑菇 (Agaricus
bisporus) 擔子果原基和擔子果轉錄基因庫 (cDNA
libraries),利用表達序列標簽法 (expressed sequence tag (EST)) 的分析均找到細胞色素
P450系統 (cytochrome P450) (Ospina-Giraldo et al.,
2000) (Agaricus express)
,當然這些基因的超級家族如何作用於子實體形態發育,則仍是未確定。
細胞色素
P450系統這個超級家族的催化許多化學反應,表現高度立體和位置的專一性 (stereo- ad
regiospecificity) ,在催化功能區結合一輔基 (prosthetic
group) -含鐵/正鐵血紅素 (heme) 給底物氧分子。而許多真菌的細胞色素
P450系統是底物誘導的 (inducible) 。
細胞色素
P450系統 (Cytochrome P450 ) 負責以下化學作用:
RH
+ NAD(P)H + H+ + O2 → ROH + NAD(P)+ + H2O
它轉化多類化學物,包括:合成麥角固醇 (ergosterol) 等激素 (steroids)、去毒作用
(detoxification) 和合成複雜的代謝物。其中麥角固醇的合成就受控於這超級家族的成員羊毛甾醇脫甲基 (lanosterol 14α-demethylase) 。
而去毒作用包括降解體外的污染物如:聚芳族碳氫化合物
(polyaromatic hydrocarbons) 、消化(assimilate)
鏈烷(alkanes)
、還原氧化氮
(Reduction of
nitrogen oxide)等,就是透過末端羥基化(Terminal hydroxylation) 鏈烷和將脂肪酸
(fatty acids) 進行ω-羥基化(ω-hydroxylation) (末端和亞末端的羥基酸
(hydroxy acids) 和二醇 (diols) 由有關的環氧化物
(epoxides) 而來)。
灰蓋鬼傘生產兩類多細胞結構:菌核
(sclerotium)和擔子果 (basidiome) ,而且兩類結構發育初期的基因是一樣的 (subroutines)。
結實基因?子實體基因?
食用和藥用大型真菌
(edible and medicinial mushrooms) 許多都是擔子菌或子囊菌,世界生產大型真菌量仍有上升的趨勢,中國是第一大生產和出口國家,但地方的平均生產量遠比許多先進國家落後,顯示在技術和品種上仍待改善和改良。世界各地使用的這類真菌大多是子實體部分,因此,研究重點都集中在找尋結實基因和子實體基因形態發育基因。
從裂褶菌 (Schizophyllum
commune) 的研究,發現:擔子果的形態發育會將細胞壁作碳源重新運用 (Wessels,
1965; Wessels & Niederpreum, 1967; Wessels & Sietsma, 1979) 。這現象在金針菇 (Flammulina
velutipes) 、鬼傘 (Coprinus congregatus) 和構巢曲霉 (Aspergillus
nidulans) 形成子實體時也有發現 (Kitamoto & Gruen, 1976;
Robert, 1977a, b; Zonneveld, 1977) 。同樣地,構巢曲霉的基因突變株因不能合成
α1→ 3 萄聚糖,而不製圓殼果 (cleistothecium) 和/或產分生孢子
(Polacheck & Rosenberger, 1977)。
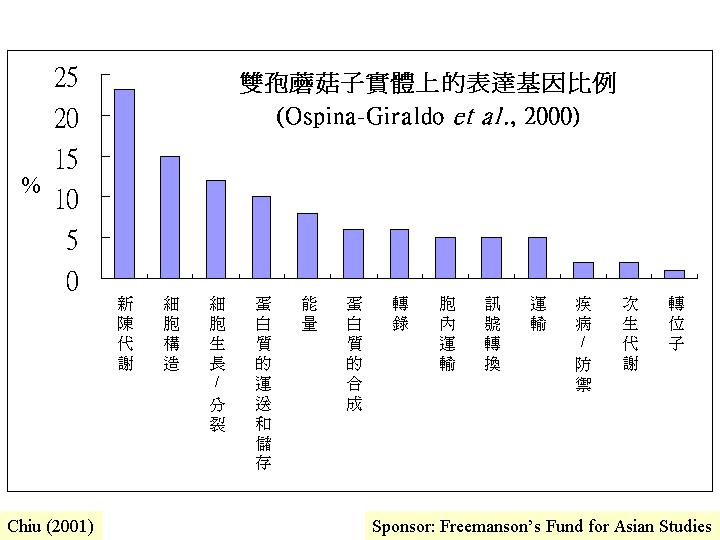
透過許多不同的生化和分子生物學技術都不難找到子實體基因 (如Opsina-Giraldo
et al., 2000) ,所謂「子實體基因」許多都指:其基因產物是在子實體大量製造,而在營養菌絲缺或沒有,但是,這些可能只代表子實體發育而增殖或擴大的基因表達,如
ATPase 或糖酵解等 (Kanda et al., 1986) ,真正與形態結構或結實或誘導形態轉向的基因,發現的太小。雙孢蘑菇子實體的表達基因中,25%與初級和次生代謝有關,25%與細胞生長、分裂和結構有關 (Agaricus
express) (Opsina-Giraldo et al., 2000)。不過,在已知的例子中,如:裂褶菌、灰蓋鬼傘和雙孢蘑菇,都確定有關子實體形態發育的基因是生物基因組
(genome) 的一小部分 (Zantinge et al., 1979;
de Vries et al., 1980; Yashar & Pukkila, 1985; Pukkila &
Casselton, 1991; Opsina-Giraldo et al., 2000)。子實體提煉出外源凝集素
(lectins) ,這類物質令(同源或外源)菌絲凝結 (Lentinula
edodes)。從裂褶菌 (Schizophyllum commune) 的研究,發現其一家族的子實體基因DNA序列編碼疏水蛋白
(hydrophobins) ,特性是:富半胱氨酸的多被分泌到體外,然後在菌絲表面聚合,當氣生菌絲露於空氣中,形成一極不溶於水的複合物 - 疏水外殼
(Wessels, 1992, 1996, 1999)。有些疏水蛋白基因 (SC1, SC3, SC4, SC7) 受控於交配型因子
(Ăsgeirsdottir et al., 1995); 單核體和雙核體都表達
SC3,疏水蛋白基因 SC1、SC4、SC6 和 SC7 只在雙核體表達,不在同交配型因子A的異核體
(common A heterokaryons) ,顯示:交配型因子 B 的開啟抑制SC3,而交配型因子A和B的親和作用激活SC1、SC4
和 SC7。免疫標記法顯示疏水蛋白SC3p只在氣生雙核體菌絲上、疏水蛋白SC7p在擔子果的細胞外基質
(extracellular matrix) 中和SC4p 包圍擔子果內的空間位置
(Wessels, 1994b)。漸漸其他真菌也可找到疏水蛋白基因,這類蛋白質負責依附在底物表面、孢子釋放和形成疏水表面等
(Hazen, 1990; Marshall, 1991; Stringer et al., 1991; Wessels et al.,
1991; Wessels, 1992, 1994; St. Leger et al., 1992; Talbot et al.,
1993; Wösten et al., 1994; Bidochka et al., 1995a, b) 。如:植病害真菌 Magnaporthe grisea 的疏水蛋白使萌發的孢子依附在稻米葉上,然後真菌才長出 appressoria引起感染
(infection) 。
真菌的疏水蛋白
(hydrophobins)。深層菌絲分泌疏水蛋白到培養基中, 疏水蛋白聚合物組織成一疏水的薄膜覆蓋培養基和降低表面張力
(surface tension), 當氣生菌絲冒出培養基時,表面就被這疏水薄膜包圍和保讓 (改自
Wessels, 1992, 2000)。菌絲接連疏水薄膜的兩個模型。 (a) 當菌絲在疏水基質表面生長而外與空氣接觸,分泌出的疏水蛋白單體
(monomer) 直接在菌絲表面上聚合, 因此, 在基質與菌絲形成一牢固的連接。(b) 假設自由的疏水蛋白單體
(monomer) 是分泌到圍繞菌絲的粘液中,經構象的改變,疏水部分結合成一不溶於SDS
(sodium dodecyl sulphate; 十二烷基磺酸鈉) 的兩性 (amphiphatic)
膜, 結果,相反的親水表面就讓菌絲接連來。注意:菌絲尖不被疏水蛋白包裸,因為聚合需要時間,而菌絲尖保持生長延伸,和分泌底物降解(Wösten
et al., 1994;
Wessels, 2000)。
疏水蛋白
(hydrophobins) 可能是真菌菌絲特有的 (Wessels, 2000)。疏水蛋白是一族小的由
100至 150胺基酸組成的分泌蛋白,DNA序列並不保守,但胺基酸序列上的保守功能區,就有8個半胱氨酸
(cysteine) 殘基,還有,整組蛋白的胺基酸鏈都表現同一模式的疏水性。疏水蛋白族可分兩組,第一組疏水蛋白
(class I hydrophobins) 可在擔子真菌找到,如: 裂褶菌 (Schizophyllum
commune) 的SC1、SC4 和 SC7 (Wessels, 2000) 和蠔菇 (Pleurotus
ostreatus) 的 POH1、POH2和POH3
(Asgeirsdottir et al., 1998)。Neurospora
crassa 的 eas基因突變種就沒有合成足夠的疏水蛋白,因而分生孢子出現水濕
(wettable) 的表現型 (Bell Pedersen et al.,
1992; Lauter et al., 1992)。
許多擔子果組織中出現各類空間 (Coprinus cinereus by John; Karen
Cheung in G. lucidum ) ( cavities ),這些可能用來提供氣體交換,但毛細管作用 (capillary action)可令空間充滿水分而喪失功能,裂褶菌 (Schizophyllum commune的 Sc4基因產物) 和雙孢蘑菇 (Agaricus bisporus) 都生產疏水蛋白 (hydrophobin) 和分泌到這些空間/ 空隙,防止貫水 (Wessels, 2000)。同樣地,地衣 (lichen) 體內有許多疏水蛋白包裸的氣間 (air spaces) 。
老化(Aging)
真菌既是單元組件生物 (modular organisms, 模祖生物、模件式的生物) 就表現不斷的生長 (Harper et al., 1986;
Andrews, 1995; Carlile, 1995) ,
生與死的觀念和動物是不一樣,
對人:死遲早會來而且可以不同形式來臨我們的身上。 Esser (1990) 指出微生物在封閉的培養下, 生命的各階段: 生長、繁殖和老化都會以: 平衡期、儲存期和維持期一一出現, 若養料能不斷供應, 真菌就處無限生長期, 但是, 單元生物上其實有不同區域的生理成熟程度, 縱是如此, 大部份的真菌會去掉一些已沒用途的組織, 如: 已散發孢子的子實體;老化子實體中, 細胞出現死亡,營養菌絲仍然存在, 可以立即或在下個適合季節結實。似乎菌絲死亡不被看作是真菌繁殖的部份 (Griffiths, 1992) , 但是在實驗環境中, 菌絲死亡是經常遇到的, 這類死亡可被視為老化, 是一漸進的失卻生長能力而致死的過程, 名菌絲老化, 而個體老化則指子實體的階段包括採摘後可儲存的時間。 細胞死亡在發育時期可有建設性功能, 其重要性與增加組織功能一樣, 同樣, 細胞死亡的時間和位置都是受控的, 因而是程式化的細胞死亡 (programmed cell death)。
柄孢殼Podospora anserina是最通常用來研究個體老化和死亡 (organismal death)現象, 因為野生品種全都表現這現象(Esser, 1990). 老化就在短暫的營養生長後發生, 因而可以量度培養的時間和接種的次量以確實其生理年齡, 不同的品種有不同的壽命, 誘發老化靠細胞核基因, 但老化這現象是核外染色體上的遺傳物質控制的;在年青細胞的線粒體上可找到質粒, 它以細胞色素氧化 (cytochrome oxidase)的亞基1(subunit 1) 中的基因內區(intron)出現, 若去除這基因內區,菌株會延長壽命的, 其機制可能是透過 RNA 切除基因內區後, 而變成一自主自我繁殖的DNA 質粒, 繼而在菌絲體散播, 感染其他年青細胞去表達老化現象 (Esser, 1990; Sainsard-Chanet
et al., 1994) 。研究結論是: 柄孢殼Podospora的老化雖然由核基因控制, 老化的誘發是一隨意的事情。同樣地, 曲霉 Aspergillus
amstelodami的營養死亡基因突變種也表現老化現象,夏威夷群島中的 Kauai島火燒地的間型孢菌自然群落中, 約30%出現老化, 現象以夏威夷語名為Kakilo (死亡的意思), Kakilo 品種的線粒體出現細胞色素短缺和多了一類DNA 質粒 (名kalDNA), 當愈多的質粒插進線粒體, 老化就發生, 而且死亡時的菌絲都很難找到線粒體DNA, kalDNA 的DNA碼根本和細胞核或線粒體的遺傳物質完全不一樣, 這便說明它是外來的, 而且能自主地在年青細胞繁殖, 插進線粒體, 而當數量過多時, 引起老化現象, 在印度作普查時, 也有找到類似Kalilo 種的粗糙孢菌種名Maranhar (Sanskrit語意死亡) (Griffiths, 1992), Maranhar種也有一鏈質粒, marDNA可自由存在或插進線粒體引起線粒體失效和老化, 這些柄孢殼和孢菌除了老化過程這病徵外, 其他生物效應則不甚清楚, 同樣地, 在自然的環境下如何發生老化也是不清楚的, 但是,這些真菌都是作為真核生物老化的研究模式,特別是: 如何控制這類質粒進入和插進適合的細胞器遺傳物質上, 而 Podospora 例子中, 因為這過程受細胞核控制, 固可視為一自我催毀的程序。
de Grey (2000) 更新一老化理論,其強綢大分子受自由基損毀引致老化和退化,線粒體 (mitochondria) 是細胞中主要受自由基攻擊的目標,同時,線粒體亦是一主要的自由基 (reactive oxygen species) 源,估計老化是長期積聚突變了線粒體的結果。線粒體的主要功能是製造 ATP,不過缺乏線粒體的細胞仍能生存,因為在胞質進行的糖酵解 (glycolysis) 仍可產生ATP,但是這些細胞的壽命是短暫的,其生存之道是靠原生質膜氧化還原的作用,降低自由基的產生
。而系統的氧化傷害是還原的 (reductive) 而非氧化的 (oxidative) 。
相反在多細胞組織的老化壽命研究中, 只集中在雙孢蘑菇上(Umar & Van Griensven,
1997b), 雙孢蘑菇的壽命在人工栽培無菌操作下約有36天的壽命,36天後的子實體細胞在電子顯微鏡下都表現退化、畸形, 而子實層上只可少量有生命的擔子。而人工栽培而生產的子實體, 老化的徵兆在18天已出現: 約50% 的子實體受木霉Trichoderma harzianum和/或 細菌Pseudomonas tolaasi 感染, 局部的細胞核和細胞質溶解,隨後細胞的滲透性增加而細胞壁有結構上的改變, 破損的痕跡在原本的位置上都可找到, 如菌柄細胞變得中空, 其他細胞缺乏支撐而倒下來, 24天後, 所在子實體都受細菌或真菌感染, 因而組織出現穿洞和腐爛。
收成的雙孢蘑菇子實體先會見到細胞壁的多處破壞, 然後是細胞質的退化,這行為被列為老化現象,但,Umar and Van Griensven (1997b)
強調自然死亡和收成的子實體老化是不一樣的。收成的子實體是因受到損害而引起死亡的, 特別是水由蒸騰作用而變得缺乏, 結果子實體表面乾旱和受損, 這其實是收割的損害, 其傷害是由外到內的, (名收割壓力引致的雜亂) ,但正常的老化 (aging) , 破壞由基因 (細胞核和細胞器的) 引起, 因而影響細胞質的整體性, 出現壞死現象 (senescence) , 是一由內致外的情況。
動物的程式化細胞死亡有兩個模式(Sen, 1992): 當細胞遇到極端非生理環境時, 細胞未能控制離子平衡, 結局是鈣跑進線粒體, 引起腫脹, 細胞質則失水, 因而不斷將環境的水份吸入, 引致細胞和細胞器過份膨脹而瓦解, 相反, 程式化的細胞死亡 (apoptotic death) 比損傷性細胞死亡 (traumatic death,
necrotic death) 較慢, 而且牽涉合成過程而引起的內退化,最後死細胞由吞噬作用而消磨。Umar and Van Griensven (1997b)
就發現雙孢蘑菇子實體有兩類細胞死亡:
菌髓的是損傷性細胞死亡
(necrotic death),
產生褶間的是真菌的程式化細胞死亡 (fungal programmed cell death)。
程式化細胞死亡的確可在灰蓋鬼傘找到 (Coprinus
PCD)。許多鬼傘種都在子實體後期出現自溶,
這是發育的一個步驟
(Buller, 1924, 1931早有這些看法), 作用是去除菌褶, 免影響擔孢子的散播。 其牽涉的在鬼傘是: 空泡(vacuole)內的几丁質、 酸性和性蛋白、RNA、磷酸 和 葡萄糖等(Iten,1968; Iten and Matile,
1970), 其實, 在營養菌絲也可找到空泡的酸性性蛋白、RNA、磷酸 和 葡萄糖活性, 因此, 這些是正常新陳代謝應用的, 但几丁質活性卻只是當擔孢子開始釋放時才製造, Iten and Matile (1970) 認為几丁質是被動地釋放的, 因為那時細胞已停止呼吸, 就如: 柄孢殼和孢菌失去線粒體活性一樣。
可見鬼傘子實體最後的自溶牽涉新的生產而且在某一特定時間發生, 這細胞死亡成為發育過程的一部分, 而且, Buller (1924)更指出, 這樣下, 菌褶會裂開 (split), 讓菌蓋如雨傘般將開, 擔孢子的釋放便落在大氣上而不是邊緣的菌褶, 因而, 緊密(close)的時空控制便是程式化細胞死亡的特徵。
Umar
and Van Griensven (1997c) 提議高級真菌在子實體發育中可能有一特別的程式化細胞死亡,如形成子實體原基時, 雖然他們都由長菌絲互纏而成, 但在極早時已可明確分清多個部位組織和平面, 這樣下, 新菌絲必須超越原有的平面而令子實體加大, 因此, 某部分的菌絲需要犧牲, 讓特定的形態去形成, 細胞學的研究在這點上都能提出支持的証據, 可見程式化細胞死亡對形成子實體扭結和原基是重要的。值得指出: 這樣的程式化細胞死亡可具有其他功能, 如鬼傘子實體死亡時有極高的溶解活性, 而細胞死後, 只留下細胞質物在體外,這些形成物(膠漿)或可提供養料作循環之用。
Goodwin (1984) 形容發育過程受基因控制就如軟件控制電腦一樣。而形態場受制於基因的有形和無形物質,包括:電、化學、機械效應等。 Trewavas (1986) 將植物發育看成代謝的網絡,代謝作用下決定組織和器官的形態,外加新元素和網絡中添加關連就啟動發育的改變,代謝的網絡就讓生物到達受外加新元素和網絡中添加關連的敏感度。真菌組織靠反饋固定作保持分化持續的模型 (feedbackfixation)展示可廣泛應用在各發育子程序的反饋固定過程,在這些發育子程序中, 週遭的微環境 (microenvironment) 與刺激物 (coactivators) 相互作用產生外加 (非遺傳)
(epigenetic) 的拱固效應,維持反饋激活環,或直接反饋固定的操控: 當細胞已定型分化發展時,外間訊號都無法改變; 減數分裂細胞 (meiocyte) 便是一例 (meiocyte)。
鳴謝:
承蒙 遠東及香港區共濟會之菲裘新研究東亞學科基金的贊助,使這部介紹真菌生理、生化、遺傳、細胞學、發育和形態及生態的資料可以中文出版,著者深致謝意! 特別鳴謝W. Bro. Peter J. Nunn 先生的支持。
The authors are deeply grateful to The Freemasons’ Fund for East Asian Studies by the District Grand Lodge of Hong Kong and the Far East, without which this publication would not have been possible. We are especially grateful to Mr. W. Bro. Peter J. Nunn, District Grand Secretary of the Fund for the kind support.
參考書目
Reference
Bőlker,
M. (2001) Ustilago maydis – a valuable model system for the study of
fungal dimorphism and virulence. Microbiology 147, 1395-1401.
de Grey, A. D. N. J. (2000)
The reductive hotspot hypothesis: an update. Archives of Biochemistry and
Biphysics 373, 295-301.
Mikosch, T. S. P., Sonnenberg,
A. S. M. & Van Griensven, L. J. L. D. (2001) Isolation, characterization
and expression patterns of a DMC1 homolog from the basidiomycete Pleurotus
ostreatus. Fungal Genetics and Biology 33, 59-66.
Muraguchi,
H. & Kamada, T. (2000) A mutation in the eln2 gene encoding a
cytochrome P450 of Coprinus cinereus affects mushroom Morphogenesis. Fungal
Genetics and Biology 29, 49-59.
Ospina-Giraldo,
M. D., Collopy, P. D., Romaine, C. P. & Royse, D. J. (2000) Classification
of sequences expressed during the primordial and basidiome stages of the
cultivated mushroom Agaricus bisporus. Fungal Genetics and Biology 29,
81-94.
Sawado, T. & Sakaguchi,
K. (1997) A DNA polymerase alpha
catalytic subunit is purified independently for the tissues at meiotic
prometaphase I of a basidiomycete, Coprinus cinereus. Biochemical and
Biophysical Research Communications 232, 454-460.
Suzuki,
K., Saga, K., Chikaoka, Y. and Itagaki, E. (1993) Purification and properties
of cytochrome P450 (P450lun) catalyzing steroid 11β-hydroxylation in Curvularia
lunata. Biochemical
and Biophysical Research Communications 263, 838-842.
Van
den Brink, H. J. M., van Gorcom, R. F. M., van den Hondel, C. A. M. J. J. &
Punt, P. J. (1998) Cytochrome P450 enzyme systems in fungi. Fungal Genetics
and Biology 23, 1-17.
Wessels
(2000) Hydrophobins, unique fungal proteins. The Mycologist 14(4),
153-159.
Please
acknowledge this homepage and authors or the source publication if you use
diagrams or materials from this website.
Thank You.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}