解讀真菌的形態發育
趙紹惠 和 大衛摩亞
Department of Biology, The Chinese University of Hong Kong, Shatin, N. T., Hong Kong SAR, China
中國香港新界沙田 香港中文大學生物系
&
School of Biological Sciences, University of Manchester, Manchester M13 9PT, U. K.
英國曼徹斯特市 曼徹斯特大學生物科學學院
第二課:真菌之生長:單細胞 (孢子/ 酵母) 及菌絲模式
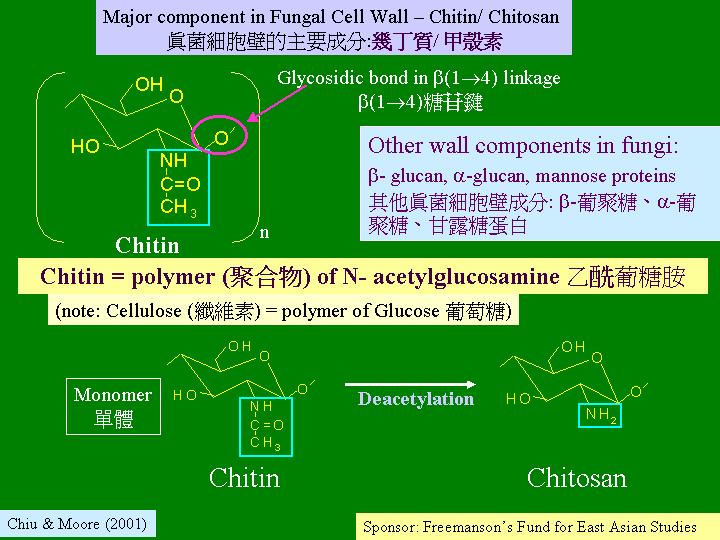
細胞壁限制細胞的形狀,細胞壁亦與形態發育有關,同時,它是環境與細胞的介面。生長和發育需要的細胞壁不能牢不可破,而是保持彈性的,隨生長、發育和環境而不斷改變,這個改變包括質和量,都是透過反應而作的。 幾丁質 (chitin) 是地球上第二多的聚合體,存在於節肢動物的外骨胳和真菌細胞壁。它和纖維素 (chitin) 相似,只是重複單位上的C-2位置不是羥基而是乙胺基 (chitin) ,多糖若有含氨基的糖或其衍生物均被稱為粘多糖 (mucopolysaccharides)。幾丁質是被幾丁質作用,水解 β-1 → 4 糖鍵而產生雙糖幾丁二糖 (chitobiose),後者被幾丁二糖轉換成單糖N-乙氨基葡萄糖 (N-acetylglucosamine;N-乙葡糖胺) ,可見幾丁質是N-乙氨基葡萄糖的聚合物 (polymer)。真菌細胞壁的合成和植物防禦病害真菌的入侵可能涉及幾丁質。真菌細胞壁不但由幾丁質 (chitin)/ 甲殼素 (chitosan) 組成,還有其他成分,包括:β-葡聚糖 (β-glucan) 和α-葡聚糖(α-glucan) 等。許多菇多糖 (mushroom polysaccharide) 就指水溶性的β-葡聚糖 (β-glucan) 或β-葡聚糖與聚合物 (β-glucan-peptide complex ) 。
極性生長
(polar growth; cytokinesis&polargrowth)
依靠指引的細胞內運輸 (directed intracellular transport)去建築和改變細胞壁,指引的細胞內運輸也影響體外的分泌。F-肌動蛋白
(F-actin)、肌球蛋白 (myosins) 和細胞骨架
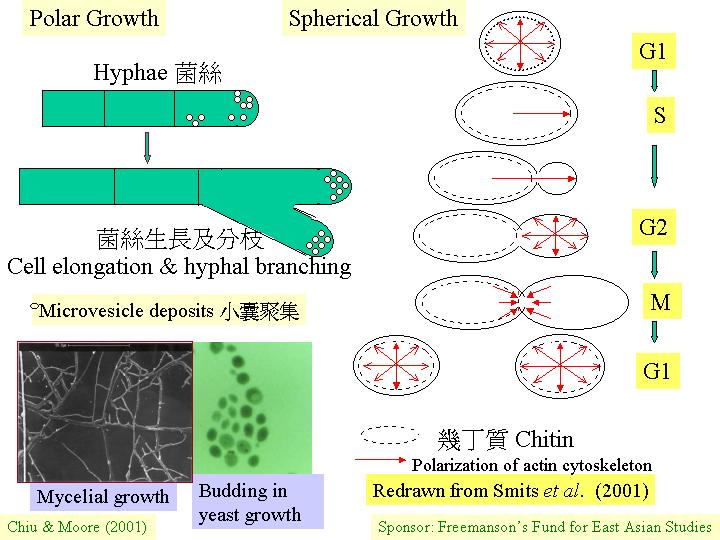
(cytoskeleton) 等均與這運輸有關。就釀酒酵母形成芽和形成(類菌絲生長模式的)交配凸出物時, 胞質成分的極性和分佈的研究發現:一束平行的微管由紡綞絲極體延伸致凸出點,若破壞微管,芽的形成不受影響,因此,微管可能只負責安放細胞核在芽領的位置和定向紡綞絲極體, 肌動蛋白的微絲則可能導向分泌泡囊到活躍的細胞壁生長處, 肌動蛋白粒在生長中的酵母是平均分佈在周邊的,但在交配凸出物則集中在頂端, 因此, 同一的細胞結構參予頂端生長和球狀生長;但不同的分佈決定兩種生長模式 (polar&sphericalgrowth)。
幾丁質(或其他降解)作用下,部分細胞壁變薄而產生不均的脹力,細胞骨架
(cytoskeleton) 因而運送和聚集合成細胞壁的超微小泡(囊)
(microvesicles) 到這位置;快速生長的粗糙孢菌 (Neurospora crassa) 管狀細胞尖上,每分鐘就約有三至四萬超微小泡與細胞膜結合
(Collinge & Trinci, 1974)。另外,相信引發真菌增殖細胞壁的訊息系統是由外至內的,其中可能與整合蛋白
(integrins) 有關;分佈在細胞膜 (cell membrane) 的整合蛋白外與細胞壁質相連而內與細胞骨架聯繫。這種細胞骨架的聯繫或可防止細胞壁發生災難性的降解。
裂殖酵母 (Schizosaccharomyces pombe)
之隔膜形成
(septum formation) 和胞質分裂 (cytokinesis) 的過程是:
(a) 細胞通過兩端延伸去生長 (稱裂殖;fission),這時,肌動蛋白
(actin) 位於細胞的生長點,而一束的微管橫跨細胞兩端,
(b) 有絲分裂 (mitosis) 開始時,肌動蛋白移位去圍繞細胞核,形成赤道絲環 (filamentous
equatorial ring),這標致將來隔膜的位置,
(c) 有絲分裂完成時,初生隔膜在周質間隙形成和向細胞內生長,
(d) 正當隔膜繼續合成時,肌動蛋白在隔膜位置由絲狀環變成簇狀小泡
(vesicles) 或粒點 (dots),
(e) 次生隔膜在初生隔膜的旁邊積聚,到成熟時,初生隔膜的降解引發細胞分裂。
與裂殖酵母形式相似的是釀酒酵母 (Saccharomyces
cerevisiae) (Smits et al., 2001),它在細胞週期 (cell
cycle) G1期時的生長是對稱的 (等方性生長,isotropic
growth),當萌芽時的芽細胞則表現不對稱生長,有絲分裂就在細胞頸進行 (即 M期),子細胞的後期生長是對稱的,到達成熟時,細胞頸原有的幾丁質隔膜就會被降解,使母與子細胞分開,重復細胞週期 (pola&sphericalgrowth)。
以上兩種酵母菌都表現:葡聚糖和幾丁質這兩個細胞壁的主要成分的合成和沉積,表現細胞週期的特性
(Smits et al., 2001) 。調控系統可能與肌動蛋白(actin) 聚合有關。
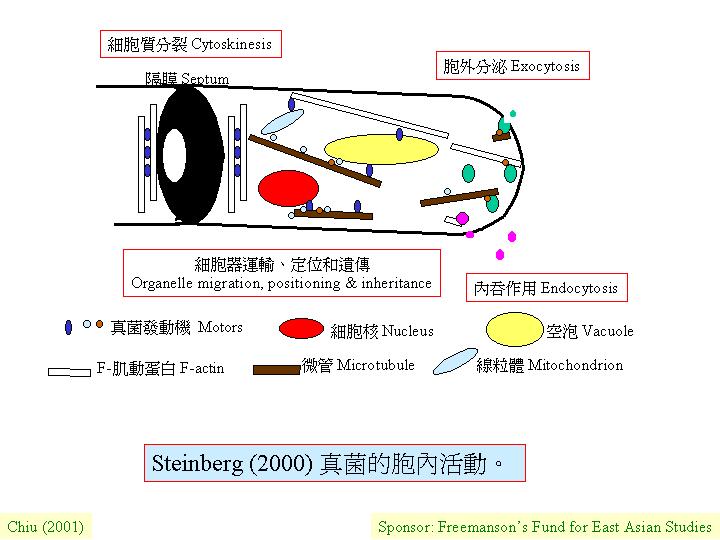
胞內運動器
(Motors)其實是大蛋白質聯合體,利用水解ATP而產生能量去推動「貨品」沿F-肌動蛋白
(F-actin) 或細胞骨架 (cytoskeleton) 路線作單一方向的
(unidirectional) 移動,所謂「貨品」包括各款細胞器 (organelles)。在結構上,胞內運動器有雙球狀功能區,各為水解ATP的反應區和一雙C-末端的功能區,作連結細胞骨架等用。F-肌動蛋白 (F-actin)
、肌球蛋白 (myosins) 和細胞骨架
(cytoskeleton) 都支持許多細胞作用,包括: 極性生長
(polar growth) 、細胞分裂/質間分裂、和有絲分裂。其實真菌有多樣策略去進片長程
(long-distance) 細胞器的運輸。
胞內運動器有三個族系
(families):
a. microtubule-based
kinesins 細胞骨架的運動蛋白
b. dyneins
(動力蛋白) (http://howard.fhcrc.org/kinesin/; http://tubulin.cb.m.u-tokyo.ac.jp/KIF/index) and
c. F-actin
(F-肌動蛋白)-associated myosins (肌球蛋白) 。
牽涉真菌胞內運動器的過程有 (cytokinesis&polargrowth):
a. 分泌
(secretion)/ 胞外分泌作用 (exocytosis) 和內吞作用
(endocytosis) 。如: 透過基因誘變和缺陷菌株的分析,在釀酒酵母 (Saccharomyces
cerevisiae) 的肌球蛋白 Myo2p是膜縛
(membrane-bound) 的, 負責運輸幾丁質合成 Chs3p (chitin synthase) 。
b. (細)胞質分裂 (Cytokinesis):
二級肌動蛋白 (class II myosin) 在細胞分裂面組結成環狀, 並與F-肌動蛋白
(F-actin) 互相作用推動過程 (cytokinesis&polargrowth)。
c. 細胞器 (organelle) 的位置、運輸和遺傳。如: 叢赤殼 (Nectria haematococa) 的kinesin的破壞,令線粒體錯放。
Kamada & colleagues
(1989a, b 和 1993) 誘導 alpha- 和 beta-微管 (tubulin) 的突變株在雙核菌絲體的核移動時發生問題,但擔子內的子核經擔孢子梗進入擔孢子的核移動則不受影響。
d. 有絲分裂
(mitosis) 。釀酒酵母 (Saccharomyces cerevisiae)
的 kinesin Kar3p在核融合時交聯在紡錘絲極體 (spindle pole
body; centrosome-equivalent organelle) 處的微管
(microtubules) ,Kar3p的存在令微管不穩定、縮短,相反,kinesin
Kip2p 和 Kip3p則穩定微管。估計 kinesins利用與極微管
(polar microtubules) 的交聯 (crosslinking) 去分隔紡綞體。dyneins (動力蛋白) 在真菌的細胞核外,故相信作用是拉(延長)星狀微管去推動核遷移或外吐小泡 (囊)
(exocytotic vesicles) 的運輸。
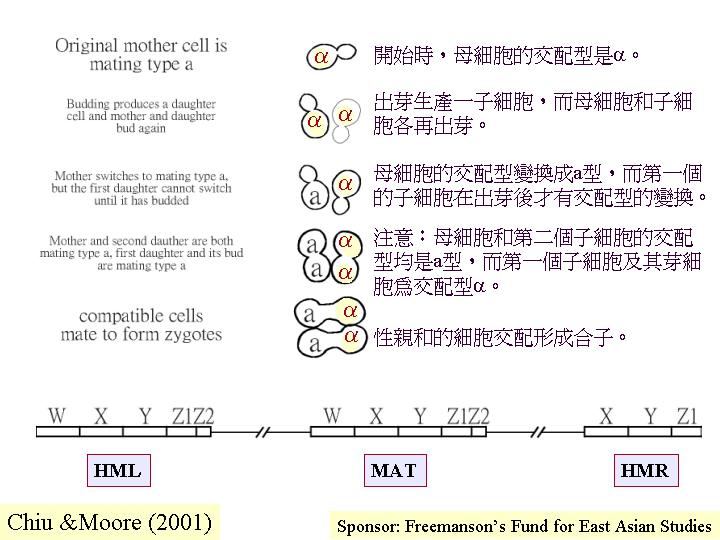
e. 基因重組
(genetic recombination) 和 RNA 運輸。釀酒酵母 (Saccharomyces
cerevisiae) 的 kinesin Kar3p 和 dyneins (動力蛋白) 的基因突變株都出現極低的減數基因重組率。而
class V myosin Myo4p就負責將調整因子 (regulator) Ash1p 的
mRNA 運輸到子細胞 (daughter cell) ,形成RNA 梯度,這調整因子是針對交配型變換 (yeastswitching專一用的內切基因的 (yeastmatingtype)。另外,擔子菌雙孢蘑菇的減數分裂後的子核分離 (nuclear segregation) 若不是隨意的,將影響後代的基因型和表現型 (Elliott &
Challen, 1983; Kerrigan et al., 1993)。
總括而言,真菌的時空組織、生長和細胞分裂都涉及胞內運動器。
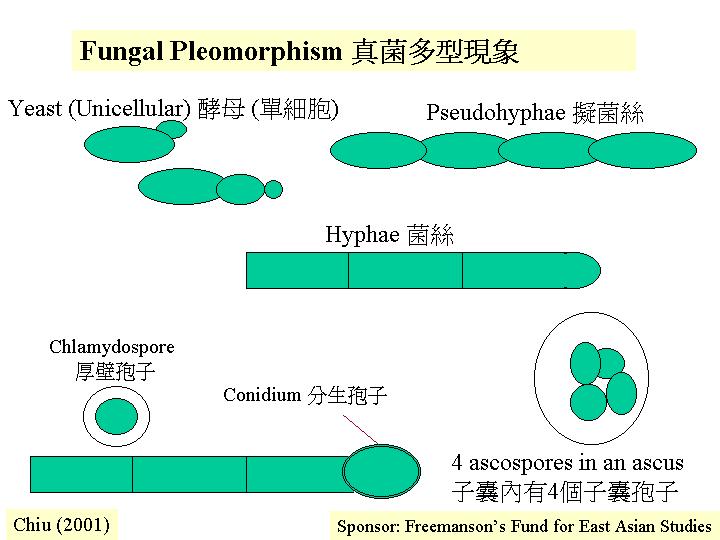
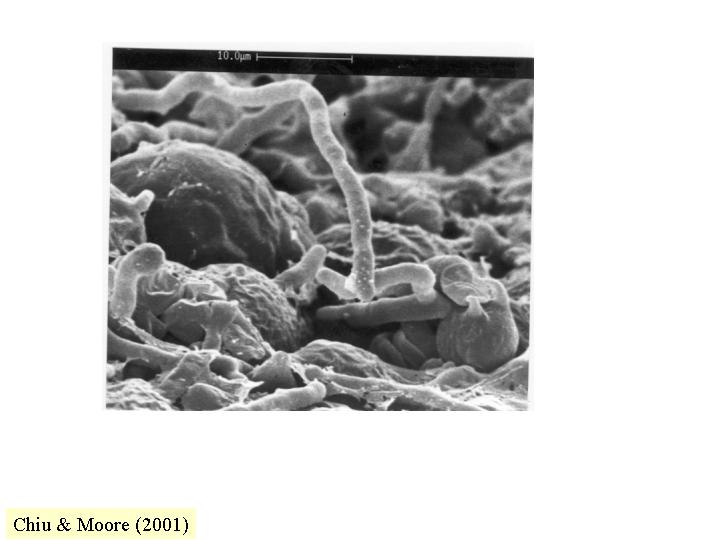
孢子、酵母與菌絲的形態與生長
白假絲酵母 (Candida albicans) 出芽
(budding) 和芽管發育時細胞壁合成的位置和形式,都表現單細胞和菌絲形式的分別。酵母
(yeast) 出芽時,細胞壁合成首先在萌芽位置出現, 在短暫的頂端延伸後, 細胞壁成輻射狀向四方八面均勻地合成, 這便是(如孢子的)球狀生長,結果, 細胞是圓形的。菌絲/假菌絲生長是持續的頂端延伸, 因此, 菌絲保持管狀 (pleiomorphism)。
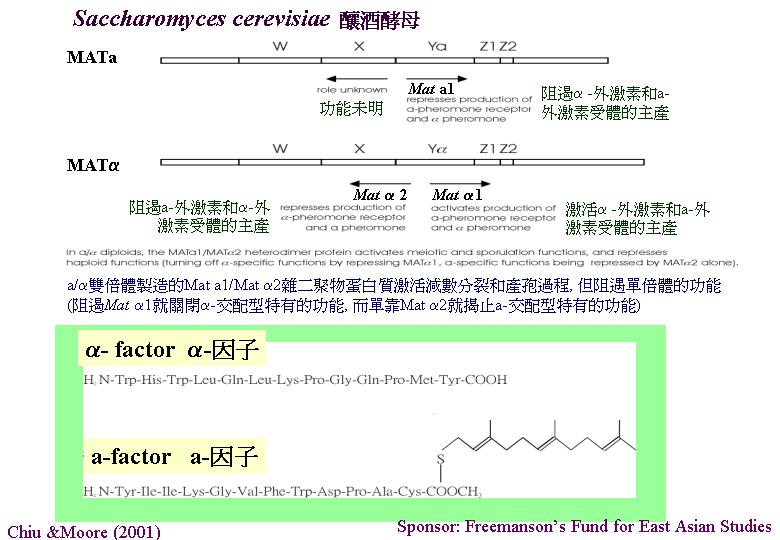
釀酒酵母 (Saccharomyces
cerevisiae) 和其他病害真菌 (黑粉菌 Ustilago
maydis、Magnaporthe grisea、白假絲酵母 (Candida
albicans) 、 和隱球酵母(Cryptococcus neoformans) 等) 都表現由單細胞轉型至假菌絲期
(pseudohypha) 。這亦是一些病害真菌由腐生
(saprobic/saprotrophic) 轉至致病階段的形態改變。當中的訊息轉導系統 (signal
transduction pathways) 由環腺酸(cAMP) 和細胞素激活的蛋白激
(mitogen-activated protein kinase) (MAPK) 調控
(Borges-Walmsley & Walmsley, 2000)。這些形態發育基因就是開發抗菌劑的目標。
饑餓或環境壓迫誘發釀酒酵母 (Saccharomyces cerevisiae) 的形態改變
在缺氮的環境下,釀酒酵母以擬菌絲形態進侵底物,有利營養的吸收。擬菌絲生長是體內G 蛋白 (如:
Ras2p) 受激活而作用的成果,這激活引發 cAMP 和細胞素激活的蛋白激
(mitogen-activated protein kinase) (MAPK) 的訊息轉導系統,連串反應包括下游的細胞表面的凝結蛋白
(flocculin) ,這細胞壁蛋白是富絲氨酸/蘇氨酸
(Ser/Thr) 的糖基化磷脂肌醇 (glycosyl phosphatidylinositol) ,負責細胞凝結作用
(flocculation)
(yeastswitching)。另外, cAMP / PKA (蛋白激 A,protein
kinase A) 路徑還調控單極萌芽,而 MAPK路徑調控細胞延伸。以下兩個程序都刺激釀酒酵母體內 cAMP 的積聚:
i.
在非醱酵性碳源(non-fermentable C-source) 培養的靜止期
(stationary phase) 細胞若接觸葡萄糖 (glucose) ,就誘發一過渡的
cAMP 峰,和
ii. 在體內酸化
(intracellular acidification) 的條件下, (如:外在環境為低 pH值時,添加氫離子(protons)
穿透劑 (protonophores) 等) ,能引起體內較高而持久的cAMP水平。
有趣地,玉米黑粉菌 (Ustilago maydis) 的腐生而非病害形態為酵母狀,而植物病害形態為菌絲狀,這形態轉換卻是經降低體內和體外的cAMP
水平而誘發的。另外,Magnaporthe
grisea的分生孢子 (conidia) 經cAMP訊息路徑
(cAMP signalling pathway) 誘發長成附胞 (appressoria),白假絲菌母 (Candida
albicans) 由萌芽過渡至菌絲/假菌絲狀都經 cAMP 訊息路徑。
擔子真菌灰蓋鬼傘 (Coprinus
cinereus) 擔子 (basidia) 的分化, 就是極性和特化的生長點, 擔孢子則表現平均生長, 這些的細胞壁添加位置就有許多高爾基小囊
(Golgi vesicles) 聚集,側絲細胞 (paraphyses) 和囊狀細胞
(cystidia) 都沒有小囊 (McLaughlin, 1974) 。Chiu
& Moore (1990) 利用一生物測試法, 考查外加的cAMP和氨離子
(ammonium) 等對擔子分化的影響,研究結果發現:cAMP 對減數分裂的擔子有抑制作用,而且導致擔子在擔孢子梗的位置長出菌絲 (圖ammoniaonbasidia.jpg
)。
其實,真菌通常是百變的,在生活史上會出現單細胞 (薄壁孢子、厚壁孢子、酵母、子囊、擔子等) 、假/真菌絲 (包括萌芽管;
pseudohypha, hypha, germ tube) 或多細胞組織等。正因為真菌可以菌絲延伸,向外擴張,一個蜜環菌 (Armillaria
gallica) 的個體有 1,500歲,面積達38 公項,而輕的孢子就可靠風高速地飄到遙遠的地方,或進入休眠期,渡過難關。 這些形態都有其功能,而形態的改變都可在細胞壁的合成/成分表達異處。
鳴謝:
承蒙遠東及香港區共濟會之菲裘新研究東亞學科基金的贊助,使這部介紹真菌生理、生化、遺傳、細胞學、發育和形態及生態的資料可以中文出版,著者深致謝意! 特別鳴謝W. Bro. Peter J. Nunn 先生的支持。
The authors are deeply grateful to The Freemasons’ Fund for East Asian Studies by the District Grand Lodge of Hong Kong and the Far East, without which this publication would not have been possible. We are especially grateful to Mr. W. Bro. Peter J. Nunn, District Grand Secretary of the Fund for the kind support.
參考書目 Key Reference
Borges-Walmsley,
M. I. & Walmsley, A. R. (2000) cAMP signalling in pathogenic fungi: control
of dimorphic switching and pathogenicity. Trends in Microbiology 8,
133-141.
Collinge, A. J. & Trinci,
A. P. J. (1974) Hyphal tips of wild type and spreading colonial mutants of Neurospora
crassa. Archives of Microbiology 99, 353-368.
Ramsdale,
M. (2001) Fungi with a sense of time: molecular genetics of temporal
organizaion in Neurospora crassa. The Mycologist 15, 10-15.
Smits,
G. J., H. van den Ende & F. M. Klis (2001) Differential regulation of cell
wall biogenesis during growth and development in yeast. Microbiology
147, 781-794.
Steinberg,
G. (2000) The cellular roles of molecular motors in fungi. Trends in
Microbiology 8, 162-168.
If
you use diagrams or material from this website, please cite the home page and
authors or the appropriate source publication in your work.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}