
4.2 Spore germination and dormancy
Spores are products of both sexual and asexual reproduction and act as the prime units of dispersal in fungi. Most spores that settle on an appropriate substrate under favourable environmental conditions germinate to produce one or more germ tubes and a new fungal mycelium. Except for sterile species, the spores of all fungi can enter into a dormant phase, during which metabolism is reduced by about 50%. Several types of dormancy exist in fungi; classed as either exogenous or endogenous (Feofilova et al., 2012). If a spore is faced with unfavourable conditions, such as lack of nutrients, low temperature, an unfavourable pH or the presence of an inhibitor (for example, on a surface of a plant), the spore remains dormant and delays germination. Spores under these conditions are exogenously dormant and will only germinate when environmental conditions become favourable.
Some fungi produce spores that fail to germinate immediately, even under favourable conditions because of factors within the spore such as nutrient impermeability or the presence of endogenous inhibitors. Spores of this sort are said to be endogenously dormant. Dormancy of these spores is usually broken by ageing or by some physiological shock permitting nutrients to begin to enter, or the endogenous inhibitors to leach out of the spore. A classic example is that dormancy of the ascospores of Neurospora crassa is broken by a 30-minute heat shock at 60°C or exposure to 0.12 mM furfural (C5H402). This relates to natural physiology because N. crassa is one of the first moulds to emerge on organic remains after bush fires. Clearly, the heat of the fire will activate dormant spores; however, furfural is prepared by acid distillation of the sugar xylose and, as many plant hemicelluloses contain xylans, bush fires are likely also to produce furfural.
Prior to the emergence of a germ tube, fungal spores undergo a process of swelling (spherical growth) during which spores increase in diameter up to four times due mainly to uptake of water. During this phase, the metabolic activity of the spore increases greatly and protein, DNA and RNA production all increase rapidly. This is followed by the emergence of one or more germ tubes (young hyphal tips) that extend away from the spore in typical apically-polarised manner (Fig. 1).
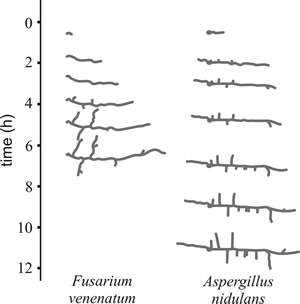 |
Fig. 1. Time lapse drawings showing development of young germlings of Fusarium venenatum and Aspergillus nidulans during the initial 12 hours of growth at 25°C on agar-solidified medium. Note that the majority of the first-formed branches are oriented at close to 90° to the long axis of the main germ tube hypha, as new hyphal tips grow directly away from their parent hypha to explore the substratum. |
Fluorescent labelling of germinating conidia of Aspergillus fumigatus (monitored by flow cytometry) revealed heterogeneities in their cell walls despite the spores having the same genetic background. The degree of heterogeneity increased during germination and was influenced by deletions of cell wall synthesising genes, environmental conditions like medium composition, method of inoculation, conidial age, and the presence of antifungal agents. The outcome of this phenotypic heterogeneity was the formation of subpopulations of germinating conidia with heterogeneous responses to the antifungal agent caspofungin, which targets cell wall synthesis, and heterogeneous sensitivities to phagocytosis by macrophages of the immune system. The phenomenon is of major clinical importance in an organism that can cause a variety of lung diseases in immunocompromised patients, including life-threatening invasive aspergillosis (Bleichrodt et al., 2020).